

Structural basis for regulation of RNA-binding proteins by phosphorylation
- PMID: 25535763
- PMCID: PMC4372107
- DOI: 10.1021/cb500860x
Structural basis for regulation of RNA-binding proteins by phosphorylation
Abstract
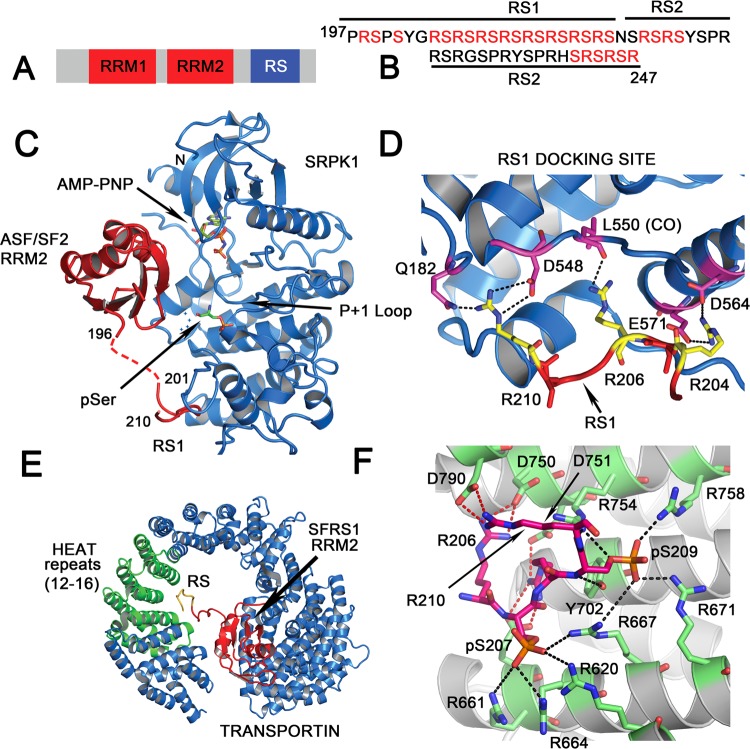
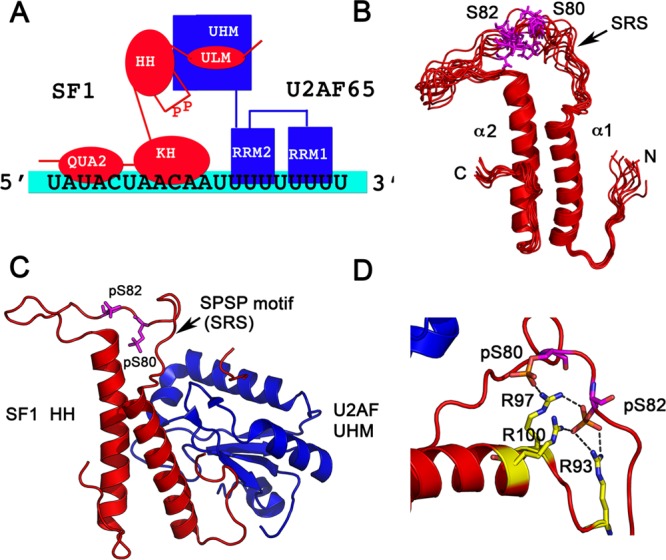
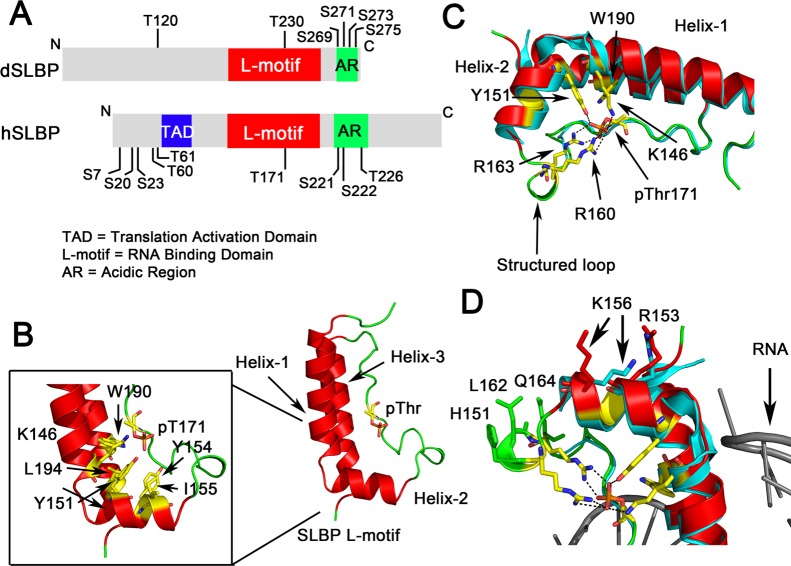
Ribonucleoprotein complexes involved in pre-mRNA splicing and mRNA decay are often regulated by phosphorylation of RNA-binding proteins. Cells use phosphorylation-dependent signaling pathways to turn on and off gene expression. Not much is known about how phosphorylation-dependent signals transmitted by exogenous factors or cell cycle checkpoints regulate RNA-mediated gene expression at the atomic level. Several human diseases are linked to an altered phosphorylation state of an RNA binding protein. Understanding the structural response to the phosphorylation "signal" and its effect on ribonucleoprotein assembly provides mechanistic understanding, as well as new information for the design of novel drugs. In this review, I highlight recent structural studies that reveal the mechanisms by which phosphorylation can regulate protein-protein and protein-RNA interactions in ribonucleoprotein complexes.
Figures

Similar articles
-
A novel protein, Pho92, has a conserved YTH domain and regulates phosphate metabolism by decreasing the mRNA stability of PHO4 in Saccharomyces cerevisiae.Biochem J. 2014 Feb 1;457(3):391-400. doi: 10.1042/BJ20130862. Biochem J. 2014. PMID: 24206186
-
Alignment of Mitotic Chromosomes in Human Cells Involves SR-Like Splicing Factors Btf and TRAP150.Int J Mol Sci. 2017 Sep 12;18(9):1956. doi: 10.3390/ijms18091956. Int J Mol Sci. 2017. PMID: 28895891 Free PMC article.
-
hnRNP A/B proteins are required for inhibition of HIV-1 pre-mRNA splicing.EMBO J. 1999 Jul 15;18(14):4060-7. doi: 10.1093/emboj/18.14.4060. EMBO J. 1999. PMID: 10406810 Free PMC article.
-
Integrated structural biology to unravel molecular mechanisms of protein-RNA recognition.Methods. 2017 Apr 15;118-119:119-136. doi: 10.1016/j.ymeth.2017.03.015. Epub 2017 Mar 16. Methods. 2017. PMID: 28315749 Review.
-
Ways and means of eukaryotic mRNA decay.Biochim Biophys Acta. 2012 Jun;1819(6):593-603. doi: 10.1016/j.bbagrm.2012.01.001. Epub 2012 Jan 12. Biochim Biophys Acta. 2012. PMID: 22266130 Review.
Cited by
-
Regulation of RNA binding proteins in trypanosomatid protozoan parasites.World J Biol Chem. 2016 Feb 26;7(1):146-57. doi: 10.4331/wjbc.v7.i1.146. World J Biol Chem. 2016. PMID: 26981203 Free PMC article. Review.
-
Lkb1 suppresses amino acid-driven gluconeogenesis in the liver.Nat Commun. 2020 Nov 30;11(1):6127. doi: 10.1038/s41467-020-19490-6. Nat Commun. 2020. PMID: 33257663 Free PMC article.
-
Arginine demethylation of Serine/Arginine-rich splicing factor 1 enhances miRNA enrichment in small extracellular vesicles derived from pancreatic ductal adenocarcinoma cells.FASEB J. 2025 Jan 15;39(1):e70287. doi: 10.1096/fj.202401811RR. FASEB J. 2025. PMID: 39760222
-
RNA stability protein ILF3 mediates cytokine-induced angiogenesis.FASEB J. 2019 Mar;33(3):3304-3316. doi: 10.1096/fj.201801315R. Epub 2018 Nov 1. FASEB J. 2019. PMID: 30383449 Free PMC article.
-
Quo Vadis Biomolecular NMR Spectroscopy?Int J Mol Sci. 2019 Mar 14;20(6):1278. doi: 10.3390/ijms20061278. Int J Mol Sci. 2019. PMID: 30875725 Free PMC article. Review.
References
-
- Nolen B.; Taylor S.; Ghosh G. (2004) Regulation of protein kinases; controlling activity through activation segment conformation. Mol. Cell 15, 661–675. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
