

Glucocorticoid regulation of lung development: lessons learned from conditional GR knockout mice
- PMID: 25535891
- PMCID: PMC5414758
- DOI: 10.1210/me.2014-1362
Glucocorticoid regulation of lung development: lessons learned from conditional GR knockout mice
Abstract
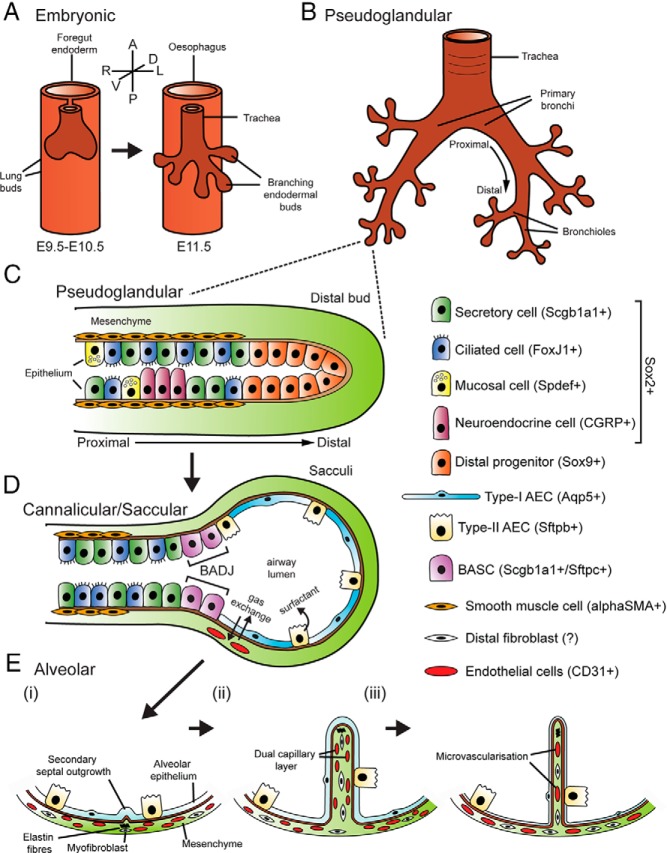
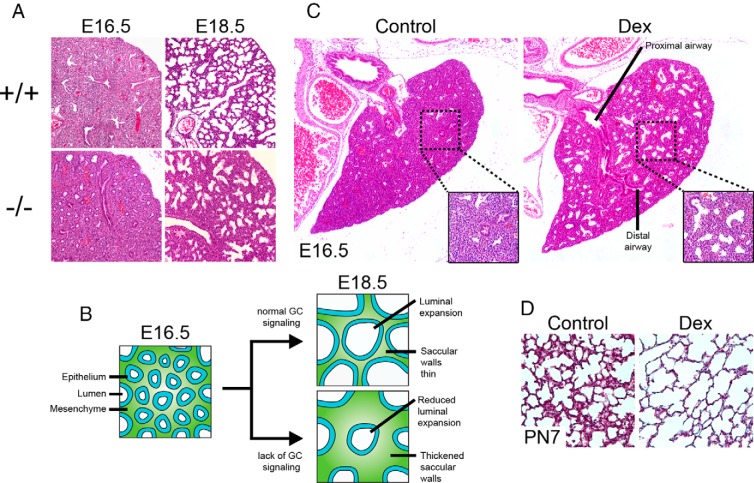
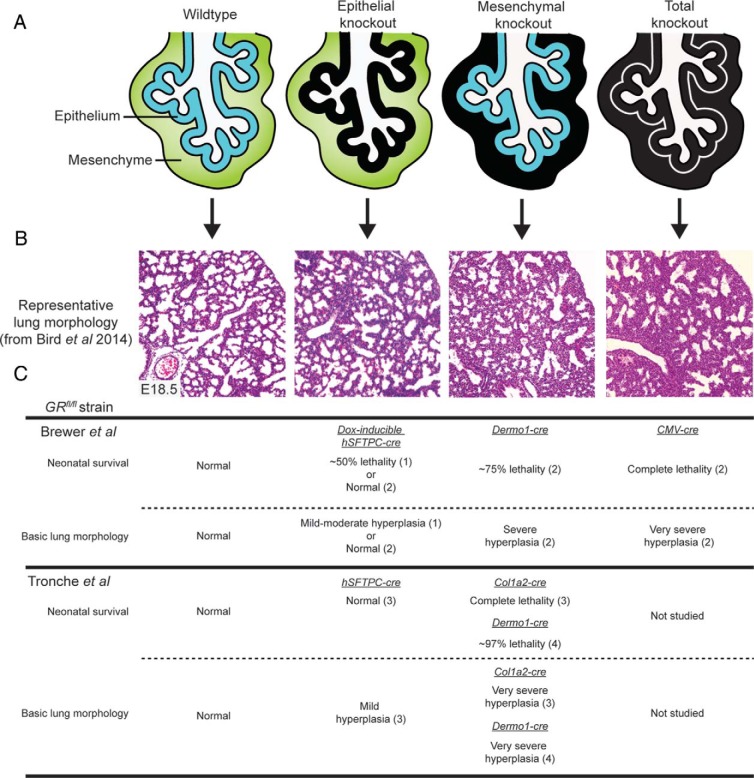
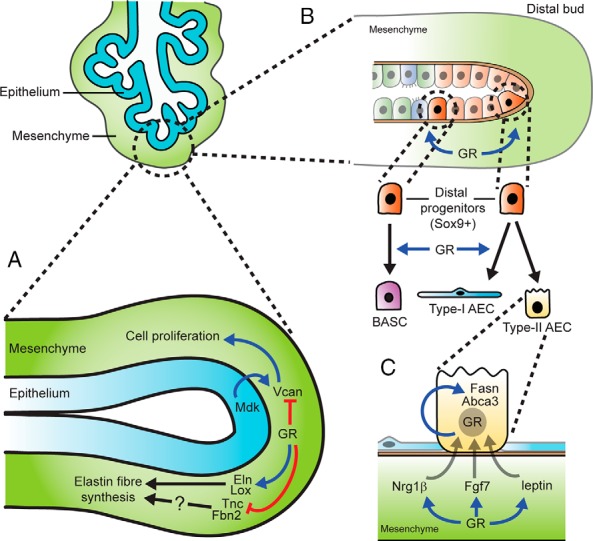
Glucocorticoid (GC) steroid hormones have well-characterized roles in the regulation of systemic homeostasis, yet less understood is their known role in utero to mature the developing respiratory system in preparation for birth. During late gestation, endogenously produced GCs thin the interstitial tissue of the lung, causing the vasculature and future airspaces to come into close alignment, allowing for efficient gas exchange at birth. More potent synthetic GCs are also used worldwide to reduce the severity of respiratory distress suffered by preterm infants; however, their clinical benefits are somewhat offset by potential detrimental long-term effects on health and development. Here, we review the recent literature studying both global and conditional gene-targeted respiratory mouse models of either GC deficiency or glucocorticoid receptor ablation. Although some discrepancies exist between these transgenic mouse strains, these models have revealed specific roles for GCs in particular tissue compartments of the developing lung and identify the mesenchyme as the critical site for glucocorticoid receptor-mediated lung maturation, particularly for the inhibition of cell proliferation and epithelial cell differentiation. Specific mesenchymal and epithelial cell-expressed gene targets that may potentially mediate the effect of GCs have also been identified in these studies and imply a GC-regulated system of cross talk between compartments during lung development. A better understanding of the specific roles of GCs in specific cell types and compartments of the fetal lung will allow the development of a new generation of selective GC ligands, enabling better therapeutic treatments with fewer side effects for lung immaturity at birth in preterm infants.
Figures
References
-
- Liggins GC, Howie RN. A controlled trial of antepartum glucocorticoid treatment for prevention of the respiratory distress syndrome in premature infants. Pediatrics. 1972;50:515–525. - PubMed
-
- Cole TJ, Blendy JA, Monaghan AP, et al. Targeted disruption of the glucocorticoid receptor gene blocks adrenergic chromaffin cell development and severely retards lung maturation. Genes Dev. 1995;9:1608–1621. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
