

CAGE-defined promoter regions of the genes implicated in Rett Syndrome
- PMID: 25539566
- PMCID: PMC4522966
- DOI: 10.1186/1471-2164-15-1177
CAGE-defined promoter regions of the genes implicated in Rett Syndrome
Abstract
Background: Mutations in three functionally diverse genes cause Rett Syndrome. Although the functions of Forkhead box G1 (FOXG1), Methyl CpG binding protein 2 (MECP2) and Cyclin-dependent kinase-like 5 (CDKL5) have been studied individually, not much is known about their relation to each other with respect to expression levels and regulatory regions. Here we analyzed data from hundreds of mouse and human samples included in the FANTOM5 project, to identify transcript initiation sites, expression levels, expression correlations and regulatory regions of the three genes.
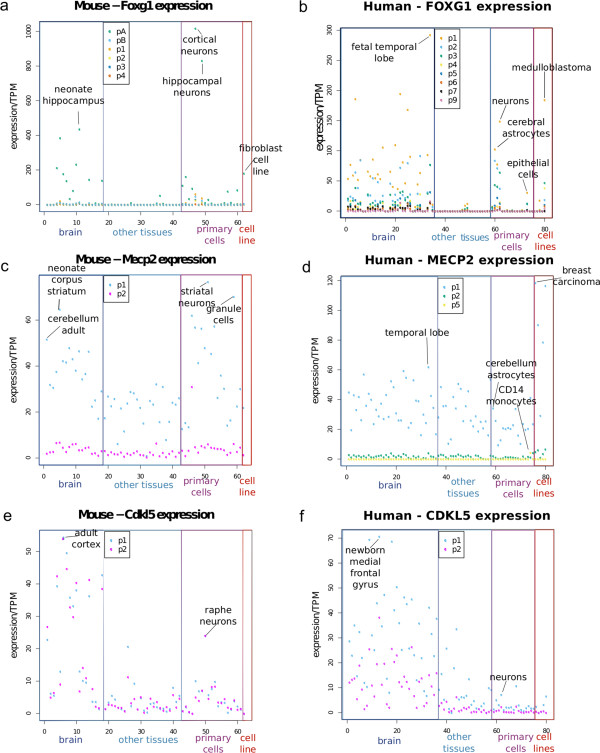
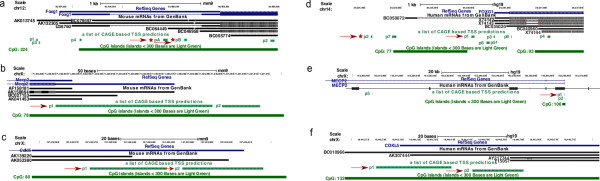
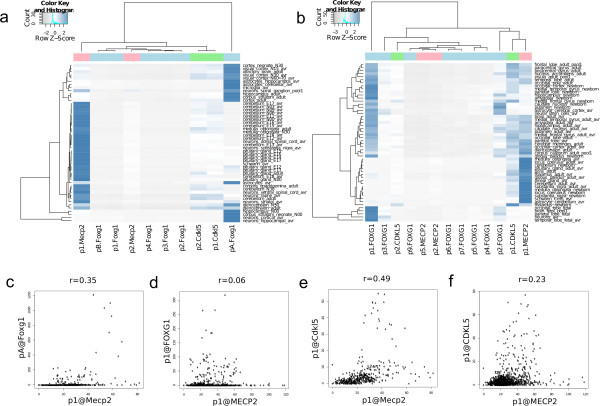
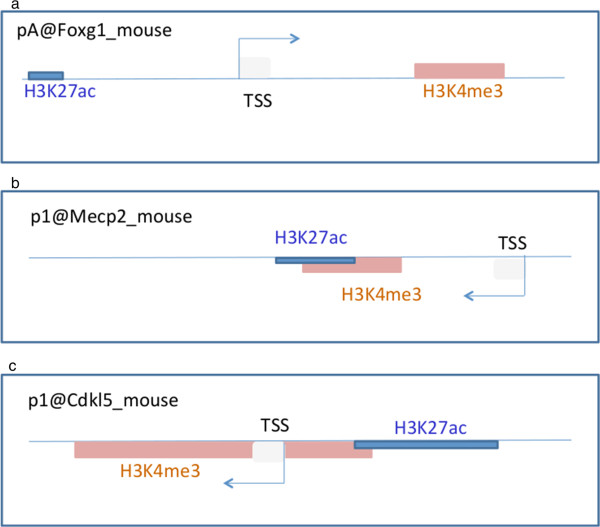
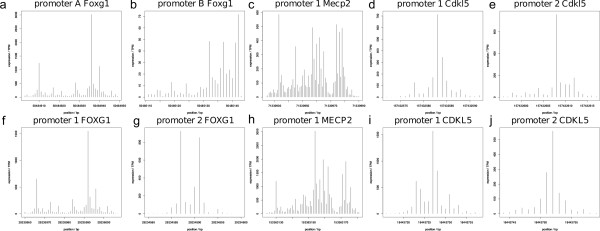
Results: Our investigations reveal the predominantly used transcription start sites (TSSs) for each gene including novel transcription start sites for FOXG1. We show that FOXG1 expression is poorly correlated with the expression of MECP2 and CDKL5. We identify promoter shapes for each TSS, the predicted location of enhancers for each gene and the common transcription factors likely to regulate the three genes. Our data imply Polycomb Repressive Complex 2 (PRC2) mediated silencing of Foxg1 in cerebellum.
Conclusions: Our analyses provide a comprehensive picture of the regulatory regions of the three genes involved in Rett Syndrome.
Figures
References
-
- Ariani F, Hayek G, Rondinella D, Artuso R, Mencarelli MA, Spanhol-Rosseto A, Pollazzon M, Buoni S, Spiga O, Ricciardi S, Meloni I, Longo I, Mari F, Broccoli V, Zappella M, Renieri A. FOXG1 is responsible for the congenital variant of Rett syndrome. Am J Hum Genet. 2008;83(1):89–93. doi: 10.1016/j.ajhg.2008.05.015. - DOI - PMC - PubMed
-
- Weaving LS, Christodoulou J, Williamson SL, Friend KL, McKenzie OL, Archer H, Evans J, Clarke A, Pelka GJ, Tam PP, Watson C, Lahooti H, Ellaway CJ, Bennetts B, Leonard H, Gecz J. Mutations of CDKL5 cause a severe neurodevelopmental disorder with infantile spasms and mental retardation. Am J Hum Genet. 2004;75(6):1079–1093. doi: 10.1086/426462. - DOI - PMC - PubMed
-
- Neul JL, Kaufmann WE, Glaze DG, Christodoulou J, Clarke AJ, Bahi-Buisson N, Leonard H, Bailey ME, Schanen NC, Zappella M, Renieri A, Huppke P, Percy AK, RettSearch Consortium Rett syndrome: revised diagnostic criteria and nomenclature. Ann Neurol. 2010;68(6):944–950. doi: 10.1002/ana.22124. - DOI - PMC - PubMed
-
- Archer HL, Evans J, Edwards S, Colley J, Newbury-Ecob R, O'Callaghan F, Huyton M, O'Regan M, Tolmie J, Sampson J, Clarke A, Osborne J. CDKL5 mutations cause infantile spasms, early onset seizures, and severe mental retardation in female patients. J Med Genet. 2006;43(9):729–734. doi: 10.1136/jmg.2006.041467. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
