

Intracellular expression of camelid single-domain antibodies specific for influenza virus nucleoprotein uncovers distinct features of its nuclear localization
- PMID: 25540369
- PMCID: PMC4325724
- DOI: 10.1128/JVI.02693-14
Intracellular expression of camelid single-domain antibodies specific for influenza virus nucleoprotein uncovers distinct features of its nuclear localization
Abstract
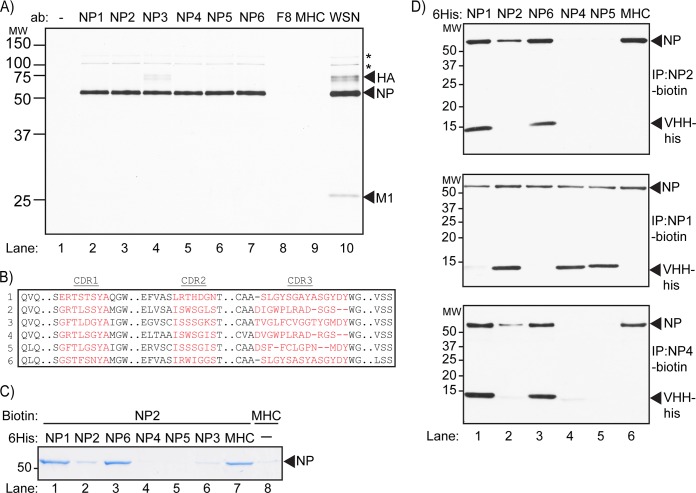
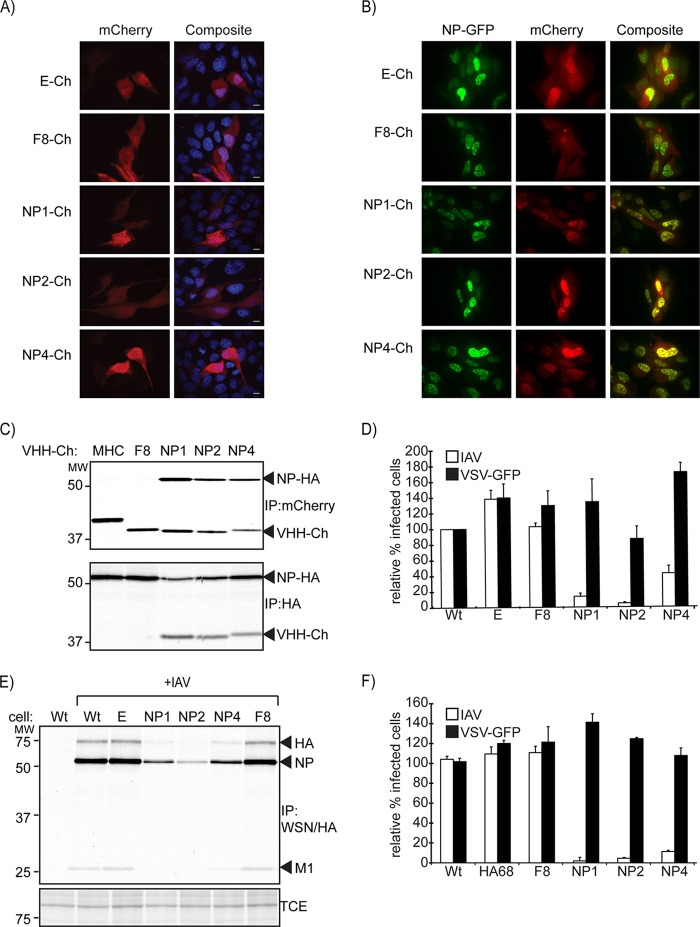
Perturbation of protein-protein interactions relies mostly on genetic approaches or on chemical inhibition. Small RNA viruses, such as influenza A virus, do not easily lend themselves to the former approach, while chemical inhibition requires that the target protein be druggable. A lack of tools thus constrains the functional analysis of influenza virus-encoded proteins. We generated a panel of camelid-derived single-domain antibody fragments (VHHs) against influenza virus nucleoprotein (NP), a viral protein essential for nuclear trafficking and packaging of the influenza virus genome. We show that these VHHs can target NP in living cells and perturb NP's function during infection. Cytosolic expression of NP-specific VHHs (αNP-VHHs) disrupts virus replication at an early stage of the life cycle. Based on their specificity, these VHHs fall into two distinct groups. Both prevent nuclear import of the viral ribonucleoprotein (vRNP) complex without disrupting nuclear import of NP alone. Different stages of the virus life cycle thus rely on distinct nuclear localization motifs of NP. Their molecular characterization may afford new means of intervention in the virus life cycle.
Importance: Many proteins encoded by RNA viruses are refractory to manipulation due to their essential role in replication. Thus, studying their function and determining how to disrupt said function through pharmaceutical intervention are difficult. We present a novel method based on single-domain-antibody technology that permits specific targeting and disruption of an essential influenza virus protein in the absence of genetic manipulation of influenza virus itself. Characterization of such interactions may help identify new targets for pharmaceutical intervention. This approach can be extended to study proteins encoded by other viral pathogens.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures


References
-
- Graham ER, Gottschalk A. 1960. Studies on mucoproteins. I. The structure of the prosthetic group of ovine submaxillary gland mucoprotein. Biochim Biophys Acta 38:513–524. - PubMed
-
- Doms RW, Helenius A, White J. 1985. Membrane fusion activity of the influenza virus hemagglutinin. The low pH-induced conformational change. J Biol Chem 260:2973–2981. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
