

Arrestins regulate cell spreading and motility via focal adhesion dynamics
- PMID: 25540425
- PMCID: PMC4325834
- DOI: 10.1091/mbc.E14-02-0740
Arrestins regulate cell spreading and motility via focal adhesion dynamics
Abstract
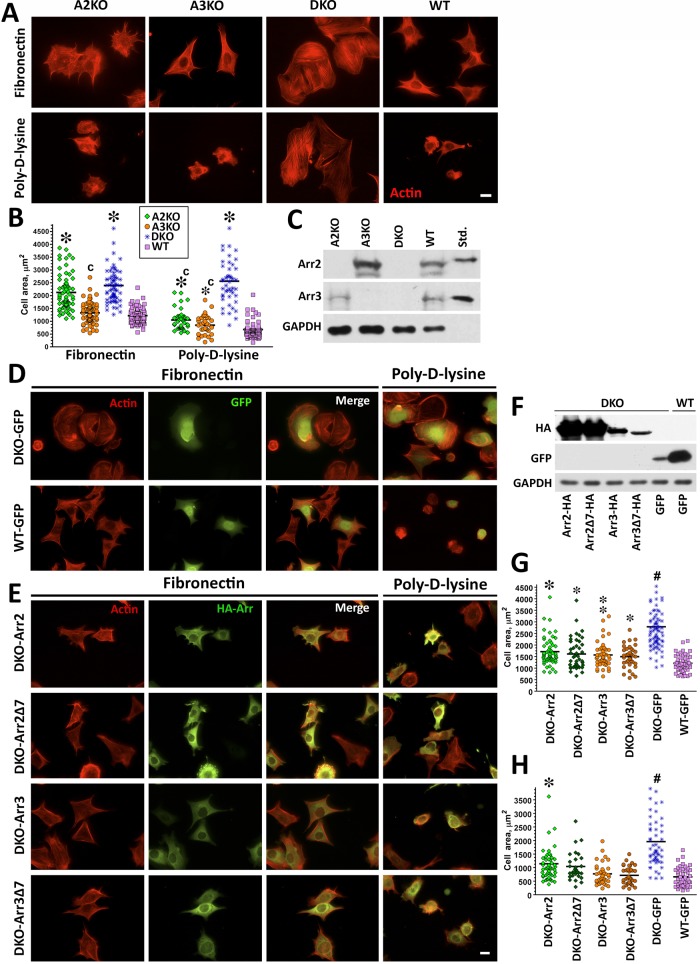
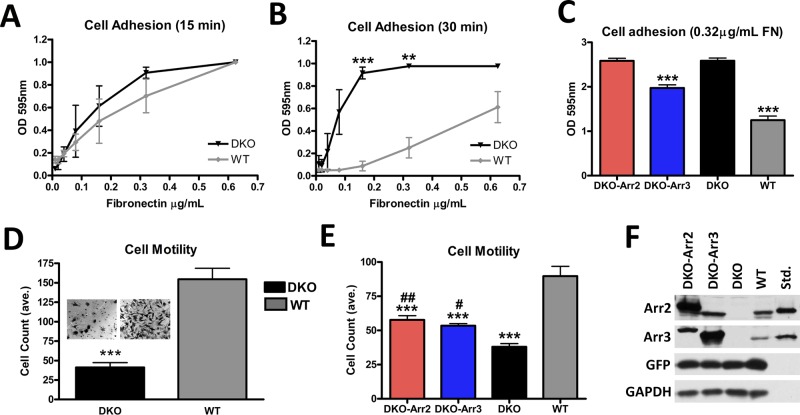
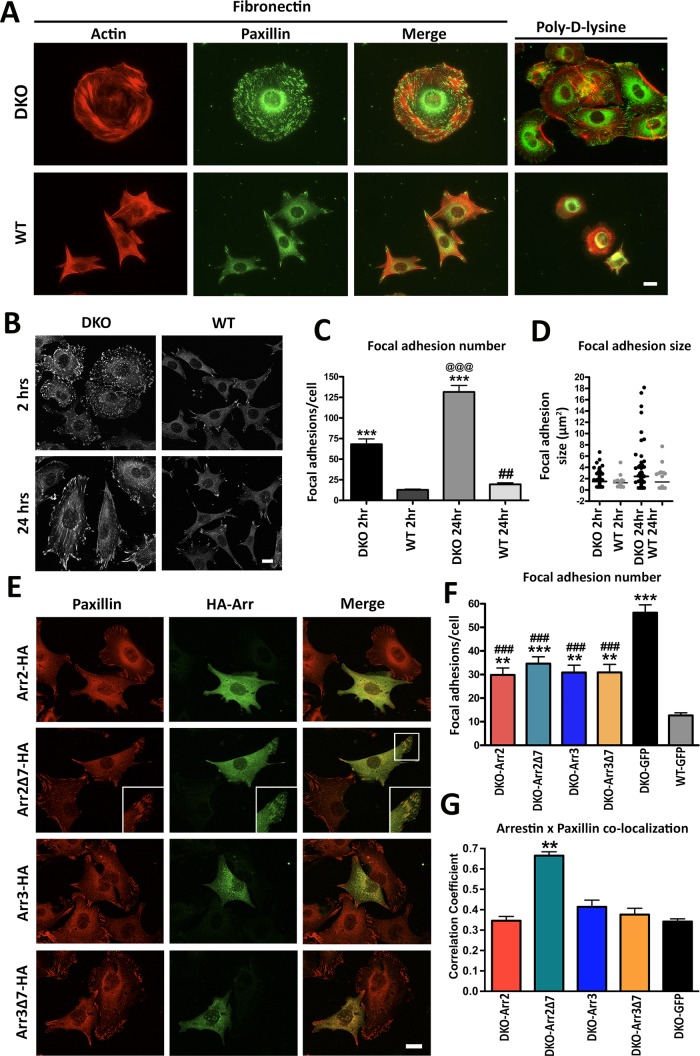
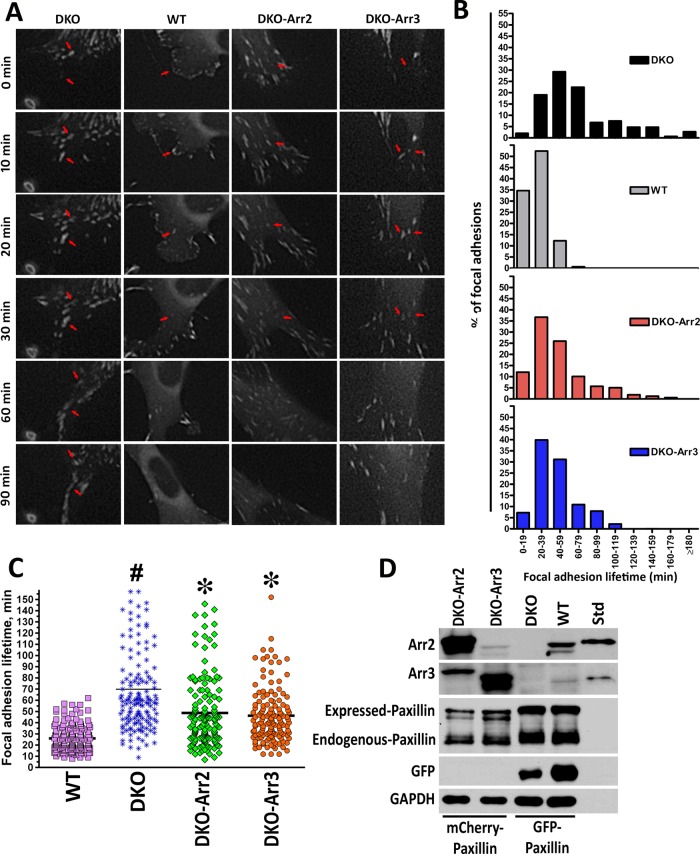
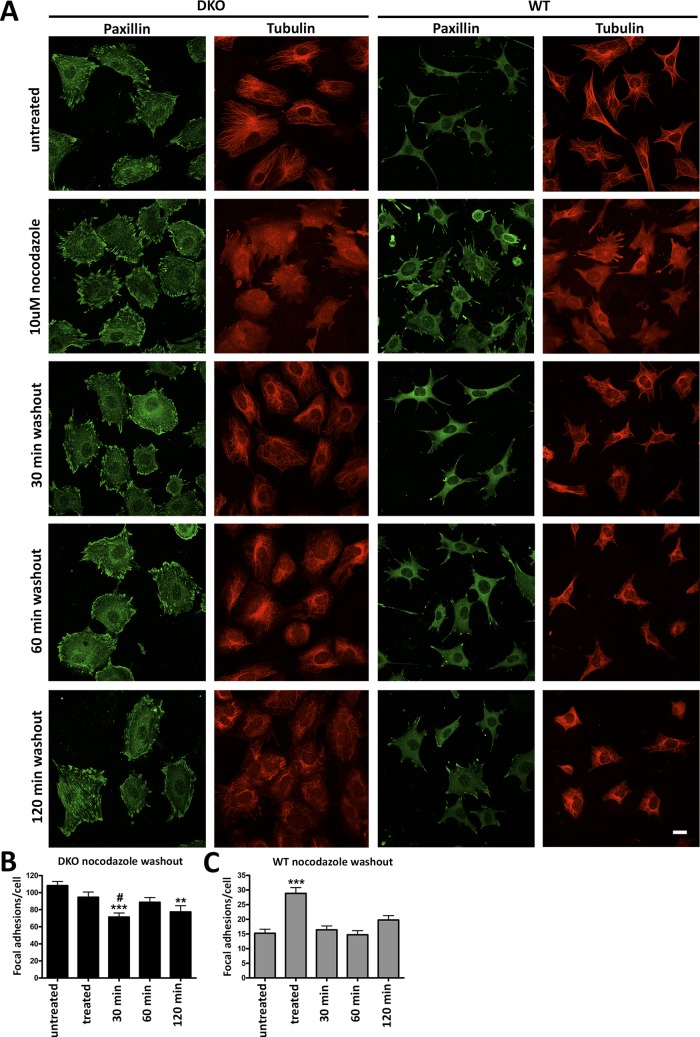
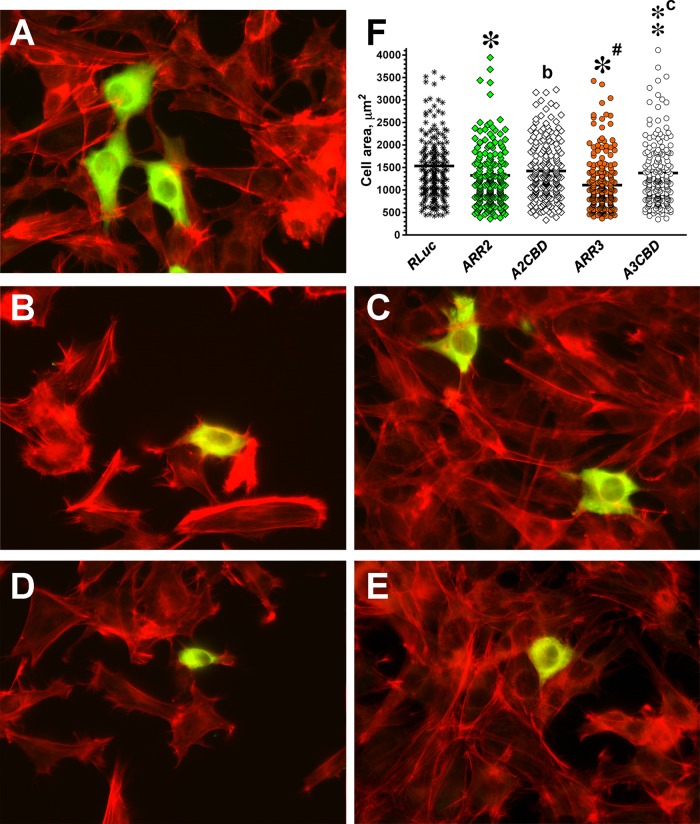
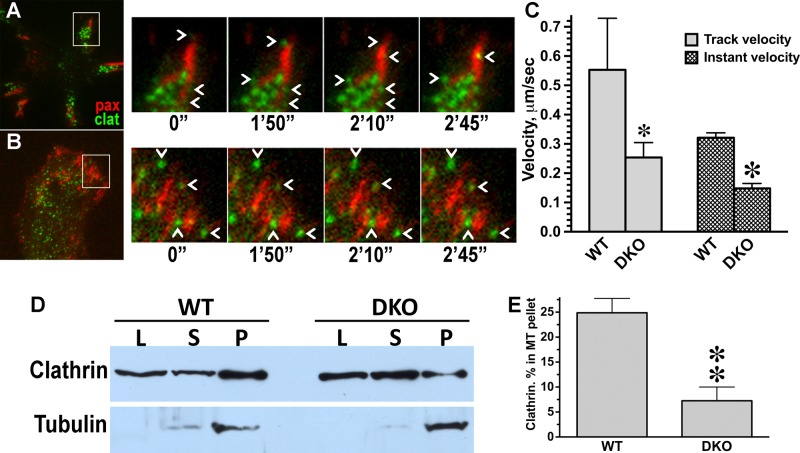
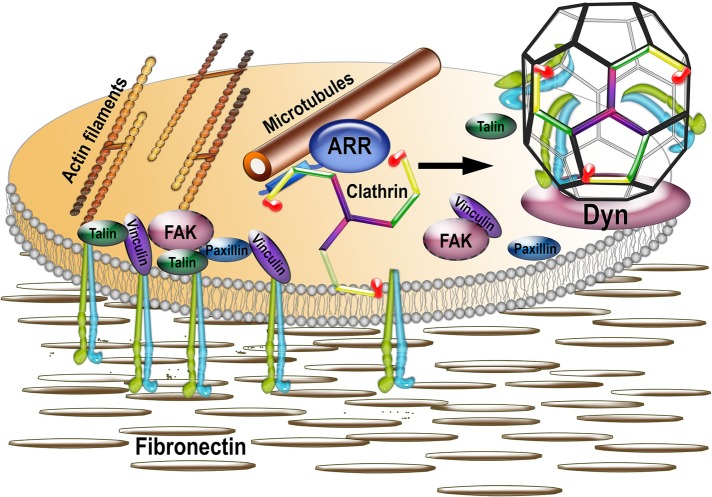
Focal adhesions (FAs) play a key role in cell attachment, and their timely disassembly is required for cell motility. Both microtubule-dependent targeting and recruitment of clathrin are critical for FA disassembly. Here we identify nonvisual arrestins as molecular links between microtubules and clathrin. Cells lacking both nonvisual arrestins showed excessive spreading on fibronectin and poly-d-lysine, increased adhesion, and reduced motility. The absence of arrestins greatly increases the size and lifespan of FAs, indicating that arrestins are necessary for rapid FA turnover. In nocodazole washout assays, FAs in arrestin-deficient cells were unresponsive to disassociation or regrowth of microtubules, suggesting that arrestins are necessary for microtubule targeting-dependent FA disassembly. Clathrin exhibited decreased dynamics near FA in arrestin-deficient cells. In contrast to wild-type arrestins, mutants deficient in clathrin binding did not rescue the phenotype. Collectively the data indicate that arrestins are key regulators of FA disassembly linking microtubules and clathrin.
© 2015 Cleghorn et al. This article is distributed by The American Society for Cell Biology under license from the author(s). Two months after publication it is available to the public under an Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).
Figures
References
-
- Anthony DF, Sin YY, Vadrevu N, Advant N, Day JP, Byrne AM, Lynch MJ, Milligan G, Houslay MD, Baillie GS. b-Arrestin 1 inhibits the GTPase-activating protein function of ARHGAP21, promoting activation of RhoA following angiotensin II type 1A receptor stimulation. Mol Cell Biol. 2011;31:1066–1075. - PMC - PubMed
-
- Barnes WG, Reiter E, Violin JD, Ren XR, Milligan G, Lefkowitz RJ. b-Arrestin 1 and Gaq/11 coordinately activate RhoA and stress fiber formation following receptor stimulation. J Biol Chem. 2004;280:8041–8050. - PubMed
-
- Bhandari D, Trejo J, Benovic JL, Marchese A. Arrestin-2 interacts with the ubiquitin-protein isopeptide ligase atrophin-interacting protein 4 and mediates endosomal sorting of the chemokine receptor CXCR4. J Biol Chem. 2007;282:36971–36979. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- T32 GM007628/GM/NIGMS NIH HHS/United States
- I01 BX002196/BX/BLRD VA/United States
- P60 DK020593/DK/NIDDK NIH HHS/United States
- CA163592/CA/NCI NIH HHS/United States
- R01 DK069921/DK/NIDDK NIH HHS/United States
- GM081756/GM/NIGMS NIH HHS/United States
- R01 CA163592/CA/NCI NIH HHS/United States
- DK58404/DK/NIDDK NIH HHS/United States
- T32 CA119925/CA/NCI NIH HHS/United States
- DA030103/DA/NIDA NIH HHS/United States
- P30 DK058404/DK/NIDDK NIH HHS/United States
- R01 GM078373/GM/NIGMS NIH HHS/United States
- R01 GM077561/GM/NIGMS NIH HHS/United States
- P30 HD015052/HD/NICHD NIH HHS/United States
- R01 GM081756/GM/NIGMS NIH HHS/United States
- NS065868/NS/NINDS NIH HHS/United States
- GM007628/GM/NIGMS NIH HHS/United States
- CA143069/CA/NCI NIH HHS/United States
- EY011500/EY/NEI NIH HHS/United States
- EY08126/EY/NEI NIH HHS/United States
- R25 GM062459/GM/NIGMS NIH HHS/United States
- EY0713516/EY/NEI NIH HHS/United States
- P30 EY008126/EY/NEI NIH HHS/United States
- R01 DK075594/DK/NIDDK NIH HHS/United States
- DK083187/DK/NIDDK NIH HHS/United States
- DK59637/DK/NIDDK NIH HHS/United States
- P30 DK020593/DK/NIDDK NIH HHS/United States
- P30 CA068485/CA/NCI NIH HHS/United States
- DK075594/DK/NIDDK NIH HHS/United States
- GM075126/GM/NIGMS NIH HHS/United States
- GM077561/GM/NIGMS NIH HHS/United States
- R21 DA030103/DA/NIDA NIH HHS/United States
- R01 DK083187/DK/NIDDK NIH HHS/United States
- R01 EY011500/EY/NEI NIH HHS/United States
- CA68485/CA/NCI NIH HHS/United States
- R01 GM075126/GM/NIGMS NIH HHS/United States
- U01 CA143069/CA/NCI NIH HHS/United States
- R01 NS065868/NS/NINDS NIH HHS/United States
- HD15052/HD/NICHD NIH HHS/United States
- DK20593/DK/NIDDK NIH HHS/United States
- GM078373/GM/NIGMS NIH HHS/United States
- DK383069221/DK/NIDDK NIH HHS/United States
- U24 DK059637/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
