

Apoplastic and intracellular plant sugars regulate developmental transitions in witches' broom disease of cacao
- PMID: 25540440
- PMCID: PMC4339597
- DOI: 10.1093/jxb/eru485
Apoplastic and intracellular plant sugars regulate developmental transitions in witches' broom disease of cacao
Abstract
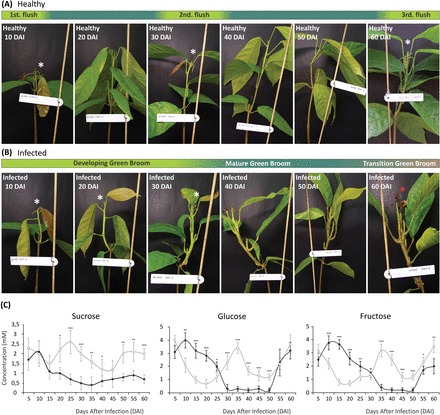
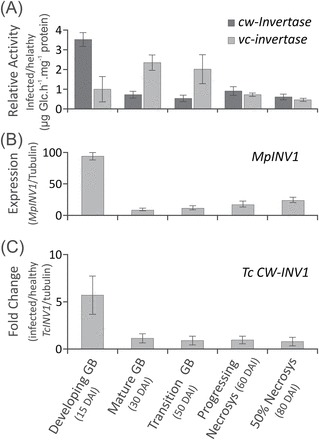
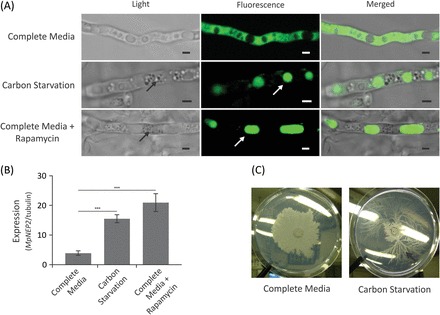
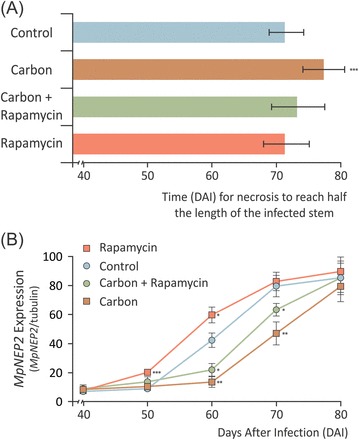
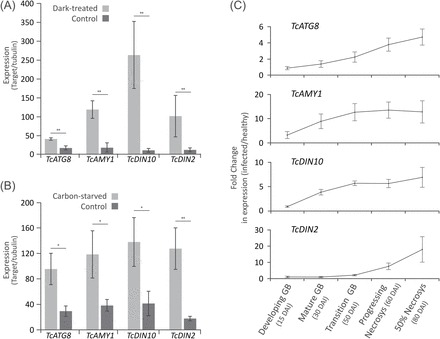
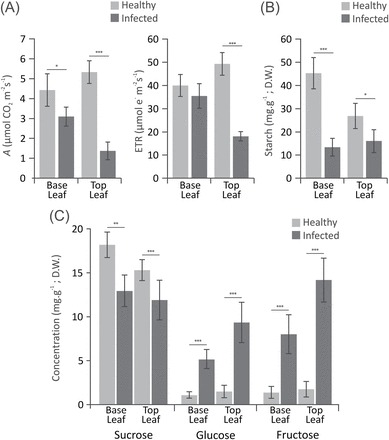
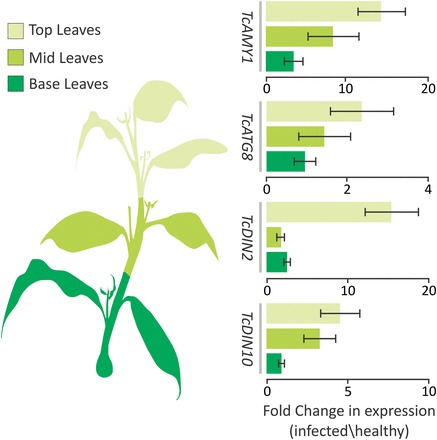
Witches' broom disease (WBD) of cacao differs from other typical hemibiotrophic plant diseases by its unusually long biotrophic phase. Plant carbon sources have been proposed to regulate WBD developmental transitions; however, nothing is known about their availability at the plant-fungus interface, the apoplastic fluid of cacao. Data are provided supporting a role for the dynamics of soluble carbon in the apoplastic fluid in prompting the end of the biotrophic phase of infection. Carbon depletion and the consequent fungal sensing of starvation were identified as key signalling factors at the apoplast. MpNEP2, a fungal effector of host necrosis, was found to be up-regulated in an autophagic-like response to carbon starvation in vitro. In addition, the in vivo artificial manipulation of carbon availability in the apoplastic fluid considerably modulated both its expression and plant necrosis rate. Strikingly, infected cacao tissues accumulated intracellular hexoses, and showed stunted photosynthesis and the up-regulation of senescence markers immediately prior to the transition to the necrotrophic phase. These opposite findings of carbon depletion and accumulation in different host cell compartments are discussed within the frame of WBD development. A model is suggested to explain phase transition as a synergic outcome of fungal-related factors released upon sensing of extracellular carbon starvation, and an early senescence of infected tissues probably triggered by intracellular sugar accumulation.
Keywords: Autophagy; Moniliophthora perniciosa; Theobroma cacao.; hemibiotrophic; senescence; starvation.
© The Author 2014. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
References
-
- Alvim FC, Mattos EM, Pirovani CP, Gramacho K, Pungartnik C, Brendel M, Cascardo JC, Vincentz M. 2009. Carbon source-induced changes in the physiology of the cacao pathogen Moniliophthora perniciosa (Basidiomycetes) affect mycelial morphology and secretion of necrosis-inducing proteins. Genetics and Molecular Research 8, 1035–1050. - PubMed
-
- Berger S, Sinha AK, Roitsch T. 2007. Plant physiology meets phytopathology: plant primary metabolism and plant–pathogen interactions. Journal of Experimental Botany 58, 4019–4026. - PubMed
-
- Bolton MD, Thomma BPHJ. 2008. The complexity of nitrogen metabolism and nitrogen-regulated gene expression in plant pathogenic fungi. Physiological and Molecular Plant Pathology 72, 104–110.
-
- Brouwer B, Ziolkowska A, Bagard M, Keech O, Gardeström P. 2012. The impact of light intensity on shade-induced leaf senescence. Plant, Cell and Environment 35, 1084–1098. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
