

Switching of pyruvate kinase isoform L to M2 promotes metabolic reprogramming in hepatocarcinogenesis
- PMID: 25541689
- PMCID: PMC4277479
- DOI: 10.1371/journal.pone.0115036
Switching of pyruvate kinase isoform L to M2 promotes metabolic reprogramming in hepatocarcinogenesis
Abstract
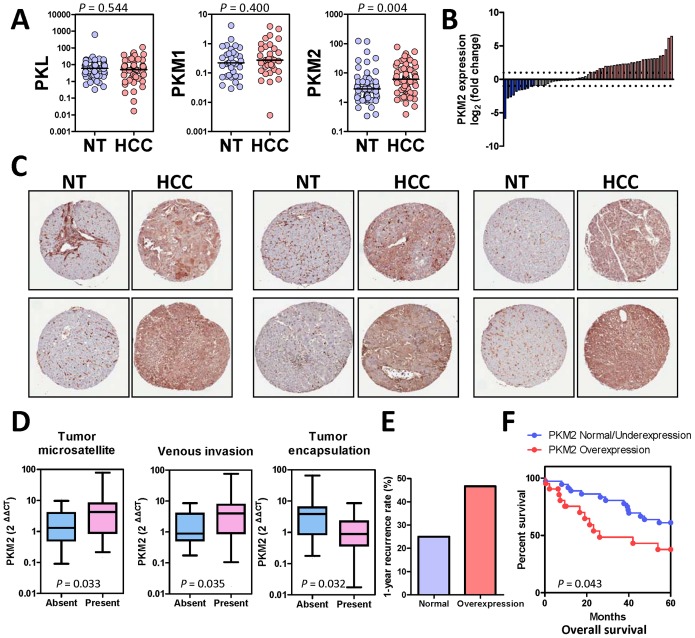
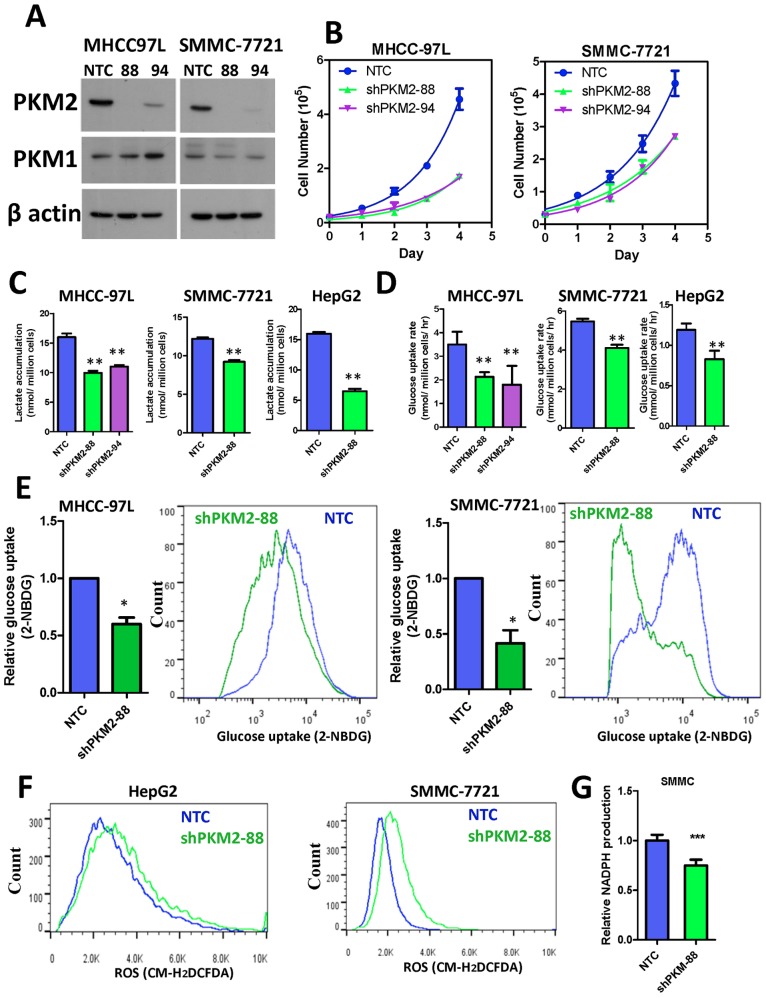
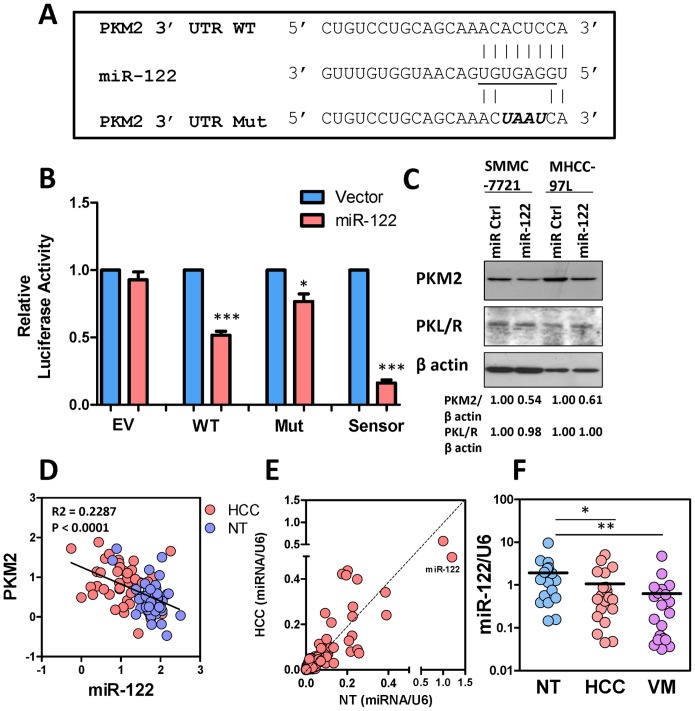
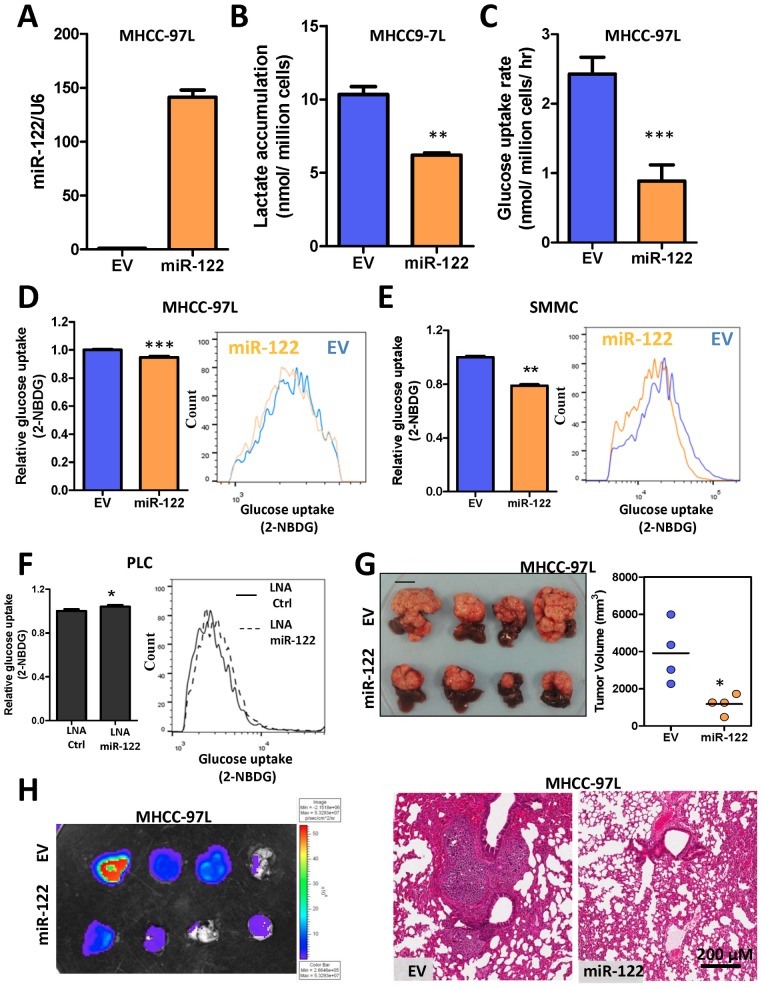
Hepatocellular carcinoma (HCC) is an aggressive tumor, with a high mortality rate due to late symptom presentation and frequent tumor recurrences and metastasis. It is also a rapidly growing tumor supported by different metabolic mechanisms; nevertheless, the biological and molecular mechanisms involved in the metabolic reprogramming in HCC are unclear. In this study, we found that pyruvate kinase M2 (PKM2) was frequently over-expressed in human HCCs and its over-expression was associated with aggressive clinicopathological features and poor prognosis of HCC patients. Furthermore, knockdown of PKM2 suppressed aerobic glycolysis and cell proliferation in HCC cell lines in vitro. Importantly, knockdown of PKM2 hampered HCC growth in both subcutaneous injection and orthotopic liver implantation models, and reduced lung metastasis in vivo. Of significance, PKM2 over-expression in human HCCs was associated with a down-regulation of a liver-specific microRNA, miR-122. We further showed that miR-122 interacted with the 3UTR of the PKM2 gene. Re-expression of miR-122 in HCC cell lines reduced PKM2 expression, decreased glucose uptake in vitro, and suppressed HCC tumor growth in vivo. Our clinical data and functional studies have revealed a novel biological mechanism involved in HCC metabolic reprogramming.
Conflict of interest statement
Figures

References
-
- Ferlay J, Shin HR, Bray F, Forman D, Mathers C, et al. (2010) Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int J Cancer 127:2893–2917. - PubMed
-
- Warburg O (1956) On the origin of cancer cells. Science 123:309–314. - PubMed
-
- Noguchi T, Yamada K, Inoue H, Matsuda T, Tanaka T (1987) The L- and R-type isozymes of rat pyruvate kinase are produced from a single gene by use of different promoters. J Biol Chem 262:14366–14371. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
