

The roles of co-chaperone CCRP/DNAJC7 in Cyp2b10 gene activation and steatosis development in mouse livers
- PMID: 25542016
- PMCID: PMC4277317
- DOI: 10.1371/journal.pone.0115663
The roles of co-chaperone CCRP/DNAJC7 in Cyp2b10 gene activation and steatosis development in mouse livers
Abstract
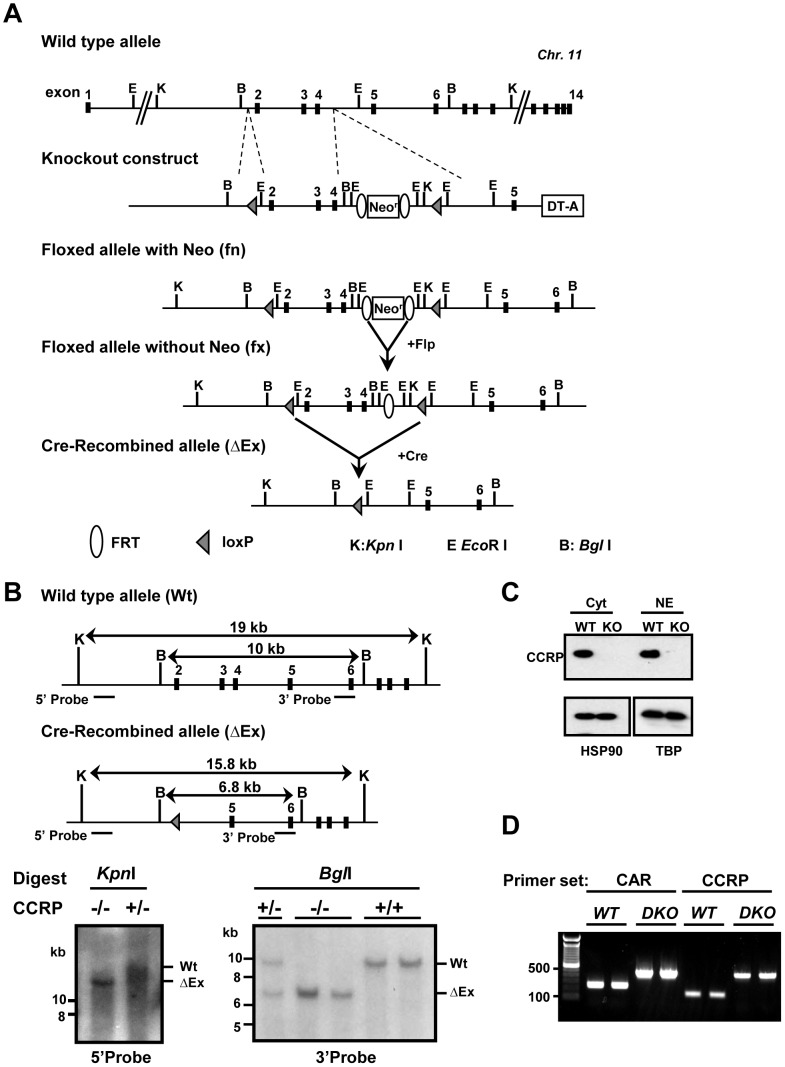
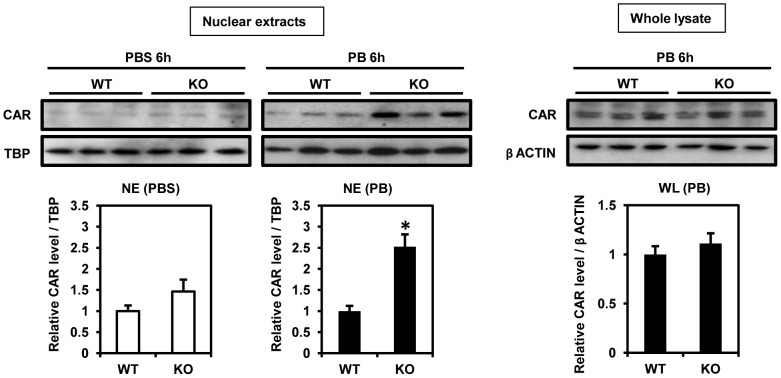
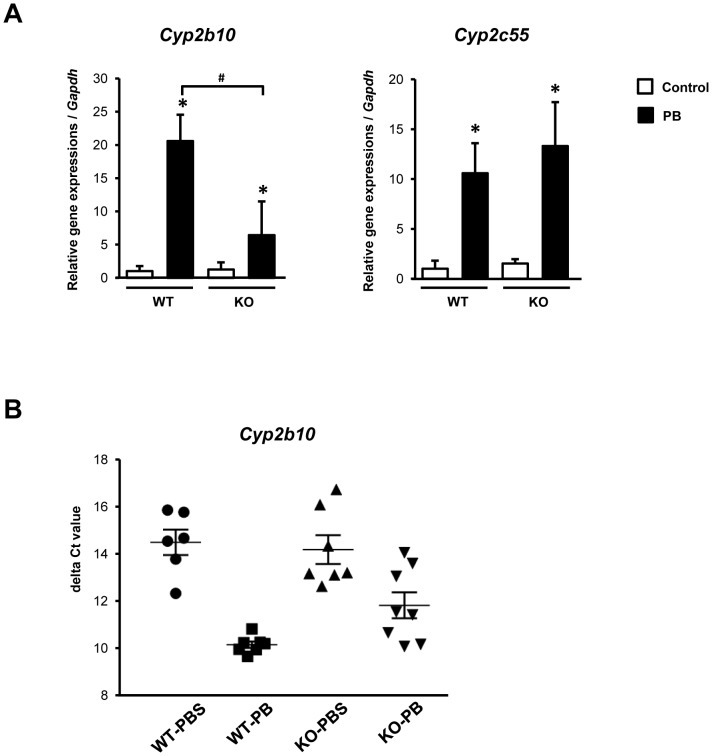
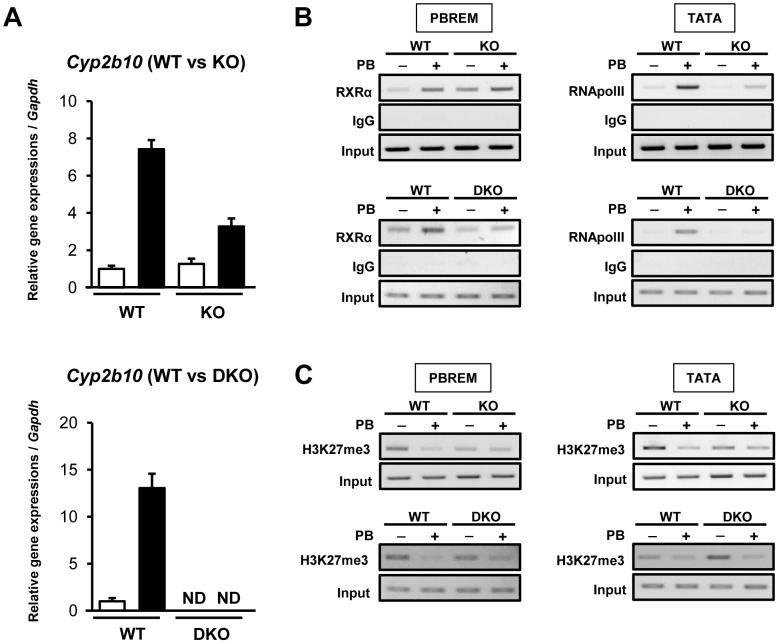
Cytoplasmic constitutive active/androstane receptor (CAR) retention protein (CCRP and also known as DNAJC7) is a co-chaperone previously characterized to retain nuclear receptor CAR in the cytoplasm of HepG2 cells. Here we have produced CCRP knockout (KO) mice and demonstrated that CCRP regulates CAR at multiple steps in activation of the cytochrome (Cyp) 2b10 gene in liver: nuclear accumulation, RNA polymerase II recruitment and epigenetic modifications. Phenobarbital treatment greatly increased nuclear CAR accumulation in the livers of KO males as compared to those of wild type (WT) males. Despite this accumulation, phenobarbital-induced activation of the Cyp2b10 gene was significantly attenuated. In ChIP assays, a CAR/retinoid X receptor-α (RXRα) heterodimer binding to the Cyp2b10 promoter was already increased before phenobarbital treatment and further pronounced after treatment. However, RNA polymerase II was barely recruited to the promoter even after phenobarbital treatment. Histone H3K27 on the Cyp2b10 promoter was de-methylated only after phenobarbital treatment in WT but was fully de-methylated before treatment in KO males. Thus, CCRP confers phenobarbital-induced de-methylation capability to the promoter as well as the phenobarbital responsiveness of recruiting RNA polymerase II, but is not responsible for the binding between CAR and its cognate sequence, phenobarbital responsive element module. In addition, KO males developed steatotic livers and increased serum levels of total cholesterol and high density lipoprotein in response to fasting. CCRP appears to be involved in various hepatic regulations far beyond CAR-mediated drug metabolism.
Conflict of interest statement
Figures

References
-
- Honkakoski P, Negishi M (1998) Regulatory DNA elements of phenobarbital-responsive cytochrome P450 CYP2B genes. J Biochem Mol Toxicol 12:3–9. - PubMed
-
- Sueyoshi T, Kawamoto T, Zelko I, Honkakoski P, Negishi M (1999) The repressed nuclear receptor CAR responds to phenobarbital in activating the human CYP2B6 gene. J Biol Chem 274:6043–6046. - PubMed
-
- Wei P, Zhang J, Egan-Hafley M, Liang S, Moore DD (2000) The nuclear receptor CAR mediates specific xenobiotic induction of drug metabolism. Nature 407:920–923. - PubMed
-
- Sueyoshi T, Negishi M (2001) Phenobarbital response elements of cytochrome P450 genes and nuclear receptors. Annu Rev Pharmacol Toxicol 41:123–143. - PubMed
-
- Sugatani J, Yamakawa K, Yoshinari K, Machida T, Takagi H, et al. (2002) Identification of a defect in the UGT1A1 gene promoter and its association with hyperbilirubinemia. Biochem Biophys Res Commun 292:492–497. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
