

Review toward all RNA structures, concisely
- PMID: 25546503
- PMCID: PMC4446244
- DOI: 10.1002/bip.22601
Review toward all RNA structures, concisely
Abstract
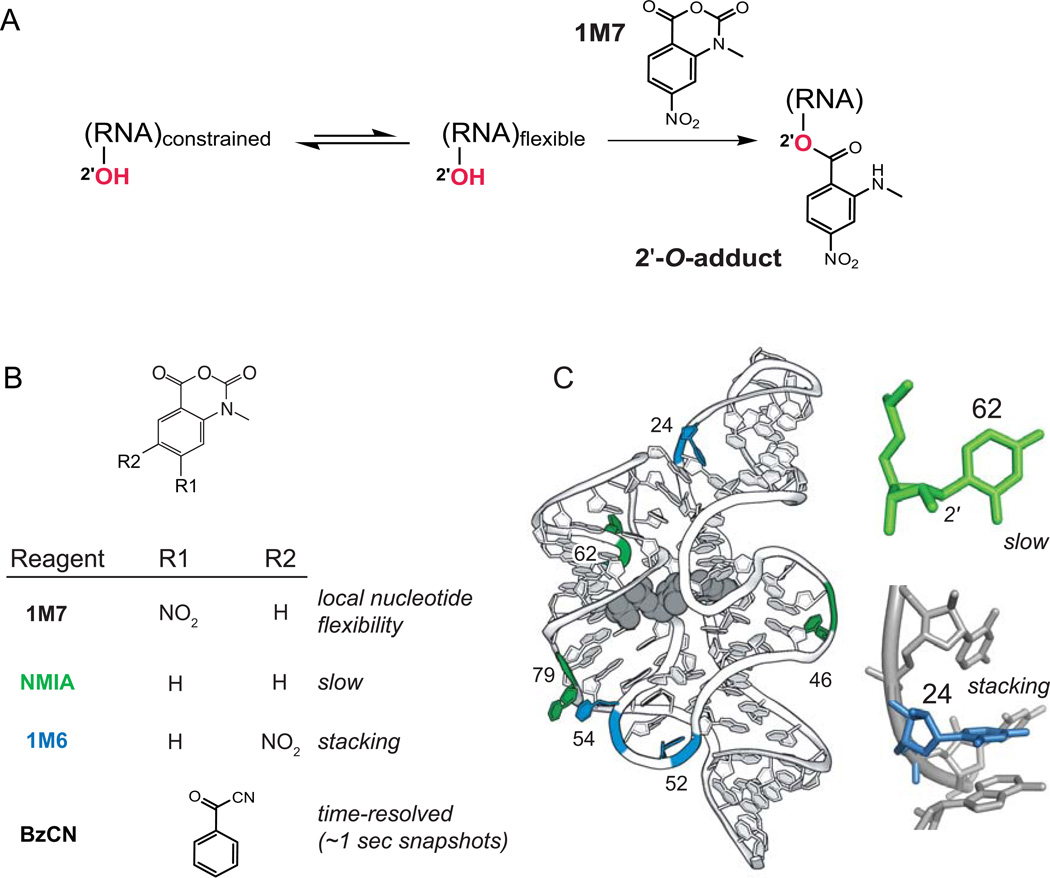
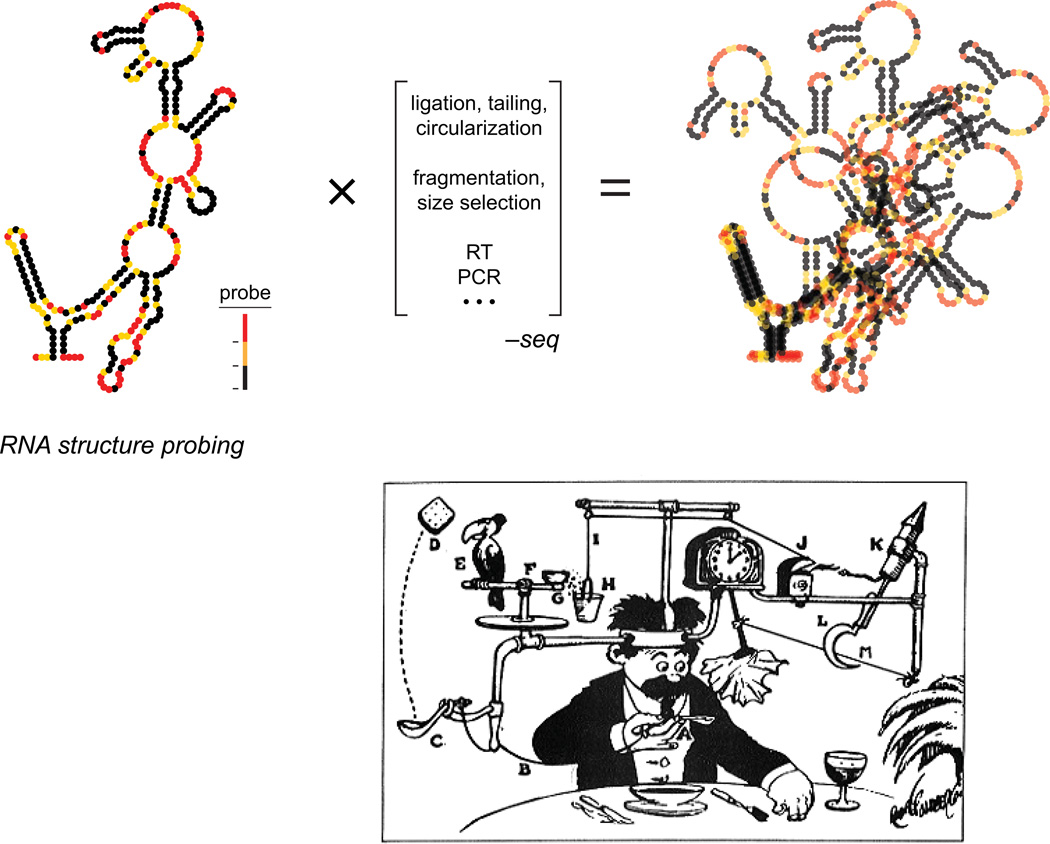
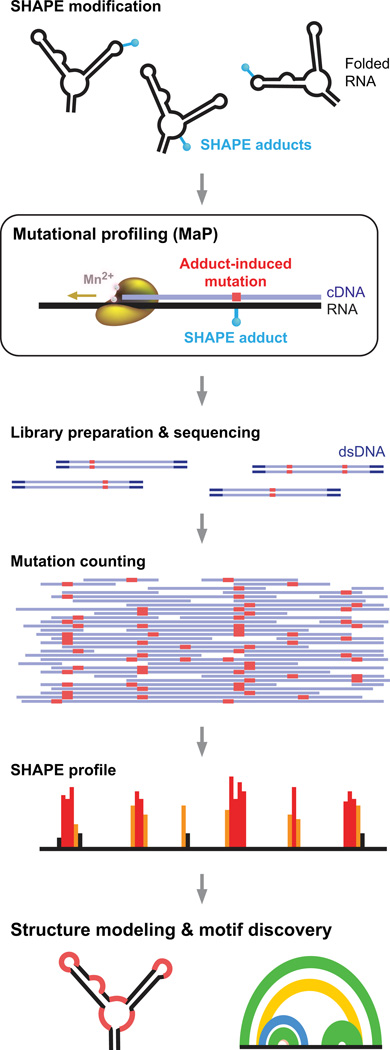
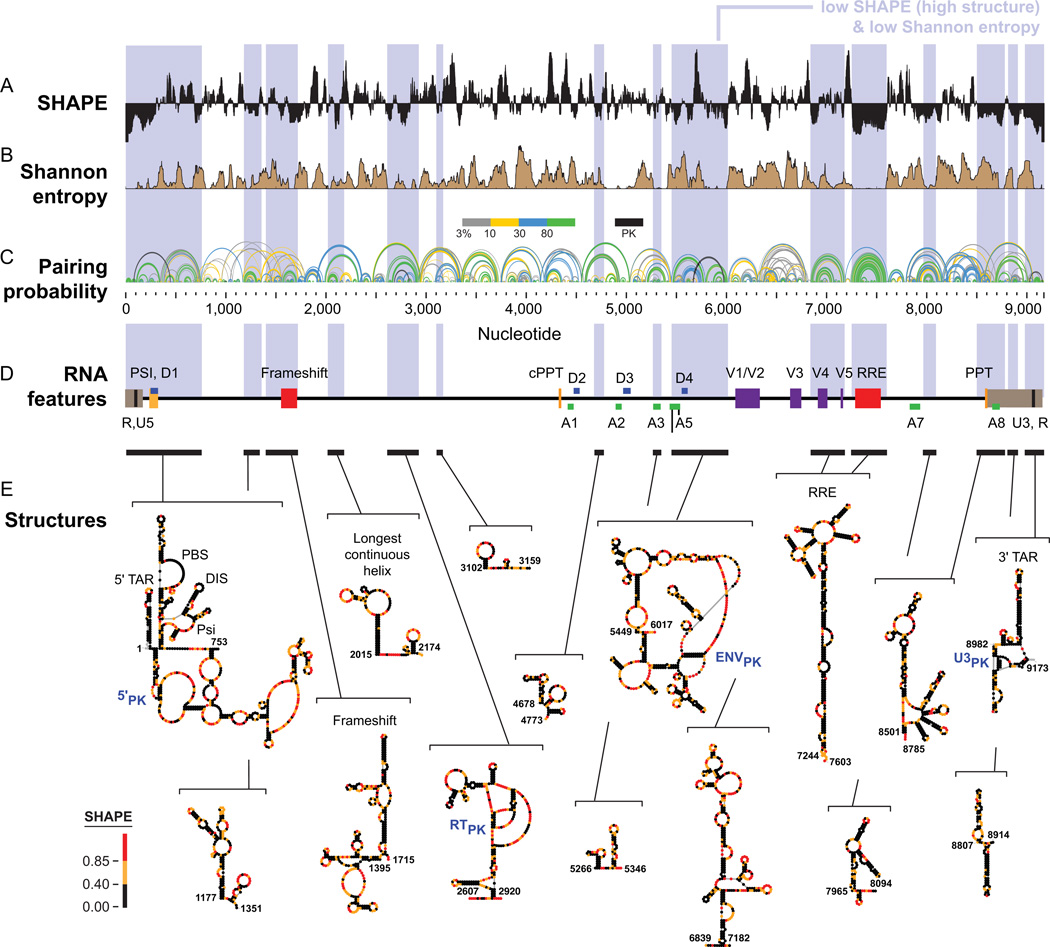
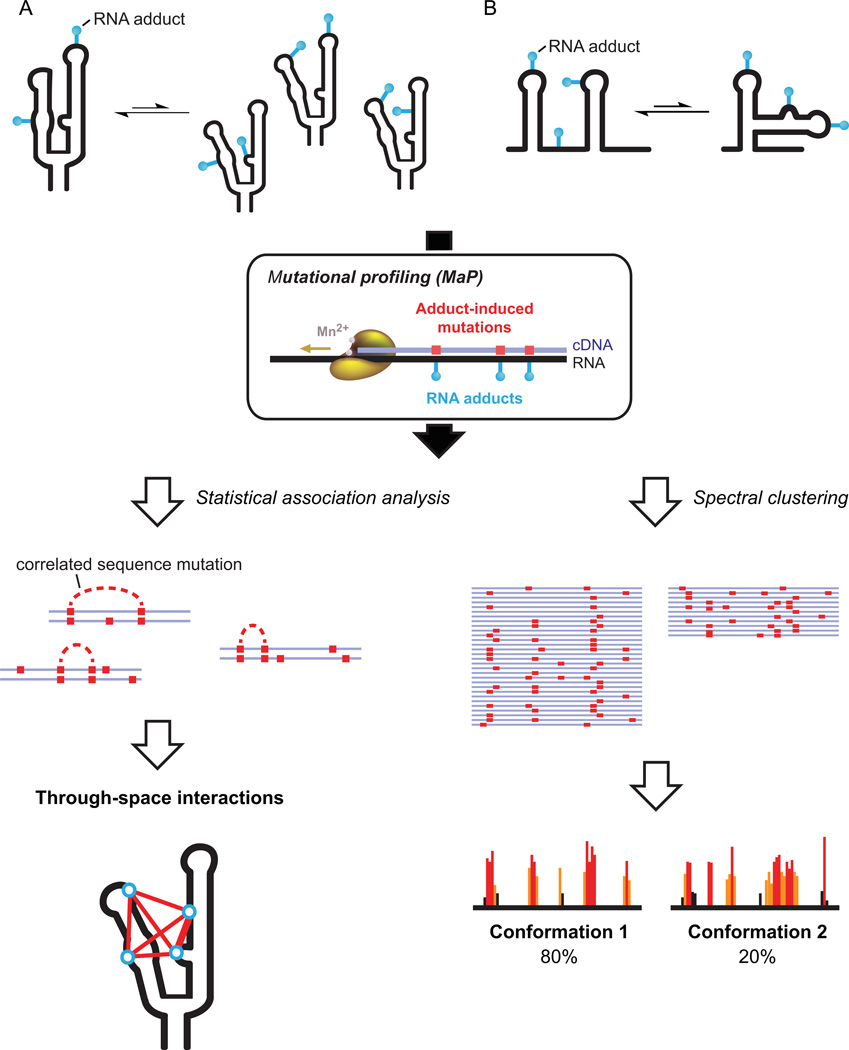
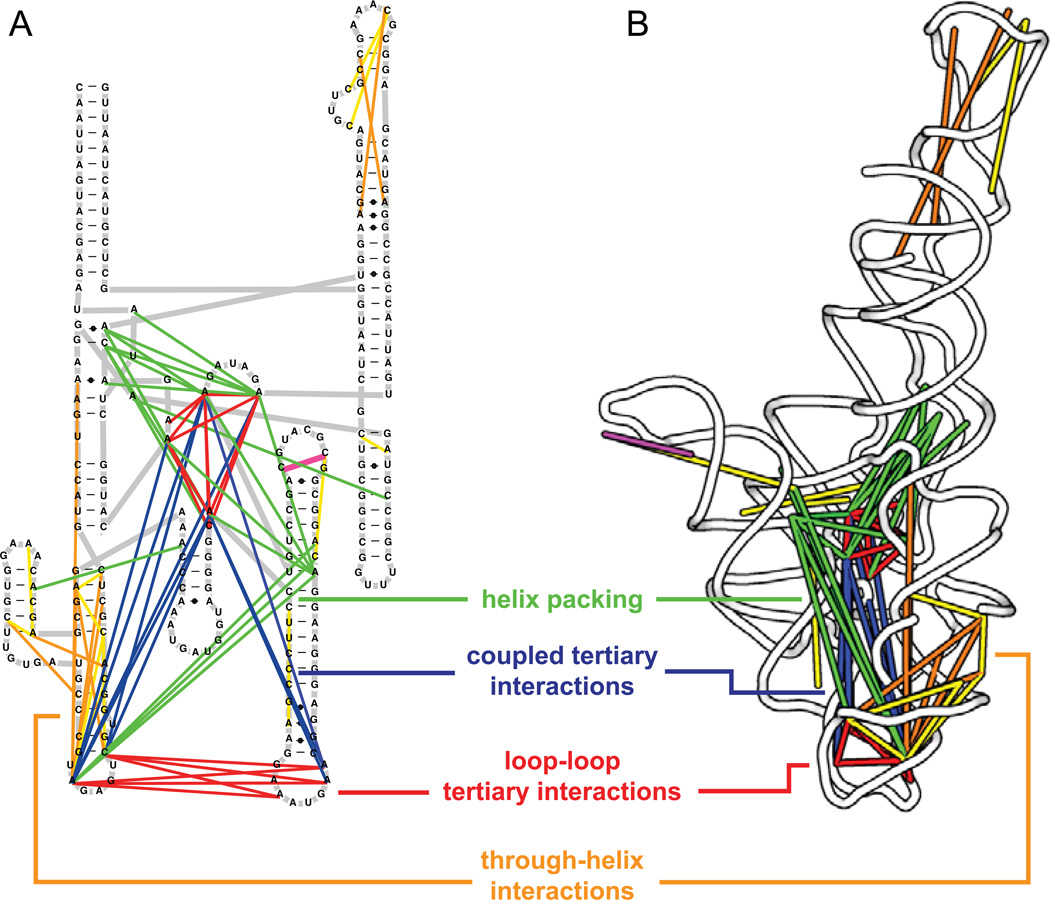
Profound insights regarding nucleic acid structure and function can be gleaned from very simple, direct, and chemistry-based strategies. Our approach strives to incorporate the elegant physical insights that Don Crothers instilled in those who trained in his laboratory. Don emphasized the advantages of focusing on direct and concise experiments even when the final objective was to understand something complex-potentially including the large-scale architectures of the genomes of RNA viruses and the transcriptomes of cells. Here, the author reviews the intellectual path, and a few detours, that led to the development of the SHAPE-MaP and RING-MaP technologies for interrogating RNA structure and function at large scales. The author also argues that greater attention to creating direct, less inferential experiments will convert "omics" investigations into lasting and definitive contributions to our understanding of biological function.
Keywords: RNA structure; chemical probing; concision; structure modeling.
© 2014 Wiley Periodicals, Inc.
Figures



References
-
-
Citation numbers were retrieved from Google Scholar on 1 Oct 2014.
-
-
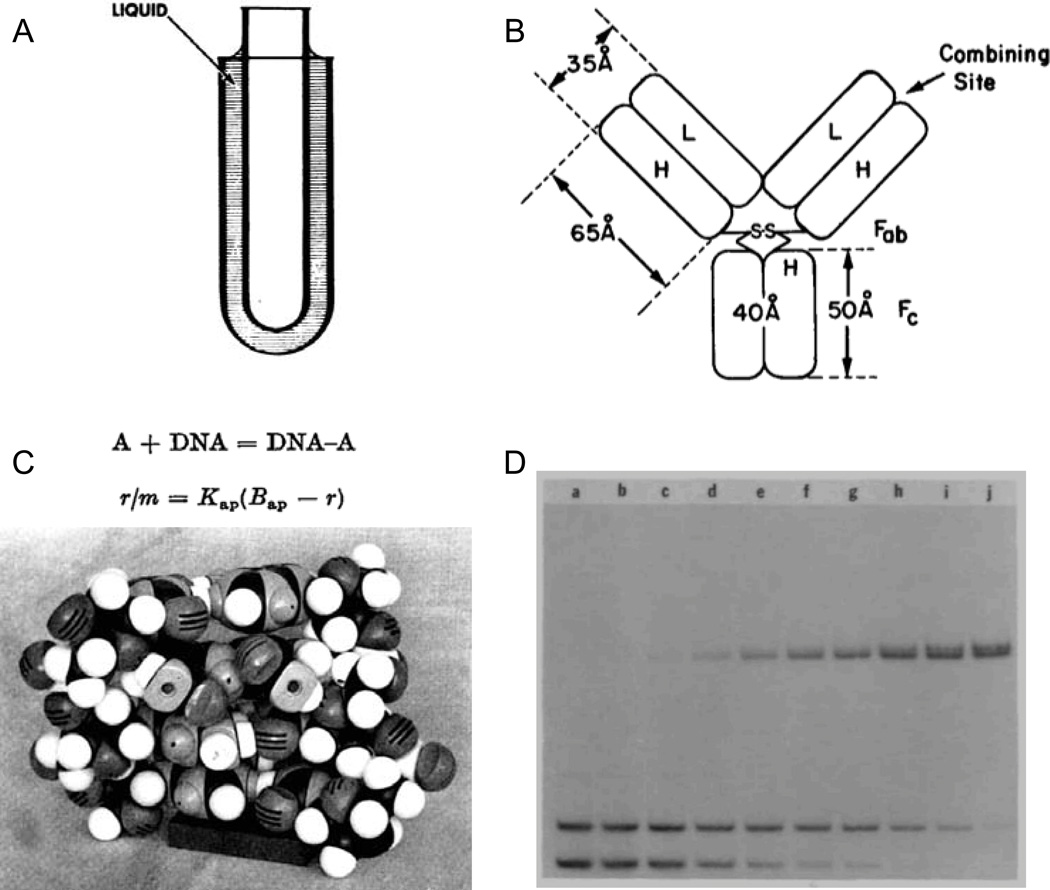
- Crothers DM, Metzger H. The influence of polyvalency on the binding properties of antibodies. Immunochemistry. 1972;9:341–357. - PubMed
-
- Müller W, Crothers DM. Studies of the binding of actinomycin and related compounds to DNA. J Mol Biol. 1968;35:251–290. - PubMed
-
- Chaires JB, Dattagupta N, Crothers DM. Studies on interaction of anthracycline antibiotics and deoxyribonucleic acid: equilibrium binding studies on interaction of daunomycin with deoxyribonucleic acid. Biochemistry. 1982;21:3933–3940. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
