

Electrochemical regulation of budding yeast polarity
- PMID: 25548923
- PMCID: PMC4280105
- DOI: 10.1371/journal.pbio.1002029
Electrochemical regulation of budding yeast polarity
Abstract
Cells are naturally surrounded by organized electrical signals in the form of local ion fluxes, membrane potential, and electric fields (EFs) at their surface. Although the contribution of electrochemical elements to cell polarity and migration is beginning to be appreciated, underlying mechanisms are not known. Here we show that an exogenous EF can orient cell polarization in budding yeast (Saccharomyces cerevisiae) cells, directing the growth of mating projections towards sites of hyperpolarized membrane potential, while directing bud emergence in the opposite direction, towards sites of depolarized potential. Using an optogenetic approach, we demonstrate that a local change in membrane potential triggered by light is sufficient to direct cell polarization. Screens for mutants with altered EF responses identify genes involved in transducing electrochemical signals to the polarity machinery. Membrane potential, which is regulated by the potassium transporter Trk1p, is required for polarity orientation during mating and EF response. Membrane potential may regulate membrane charges through negatively charged phosphatidylserines (PSs), which act to position the Cdc42p-based polarity machinery. These studies thus define an electrochemical pathway that directs the orientation of cell polarization.
Conflict of interest statement
The authors have declared that no competing interests exist.
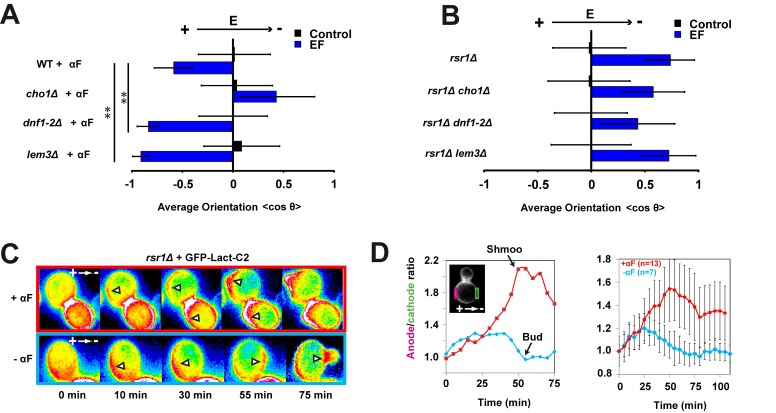
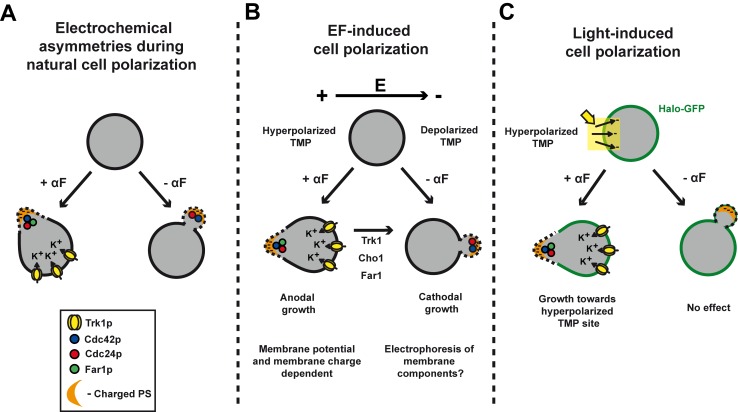
Figures
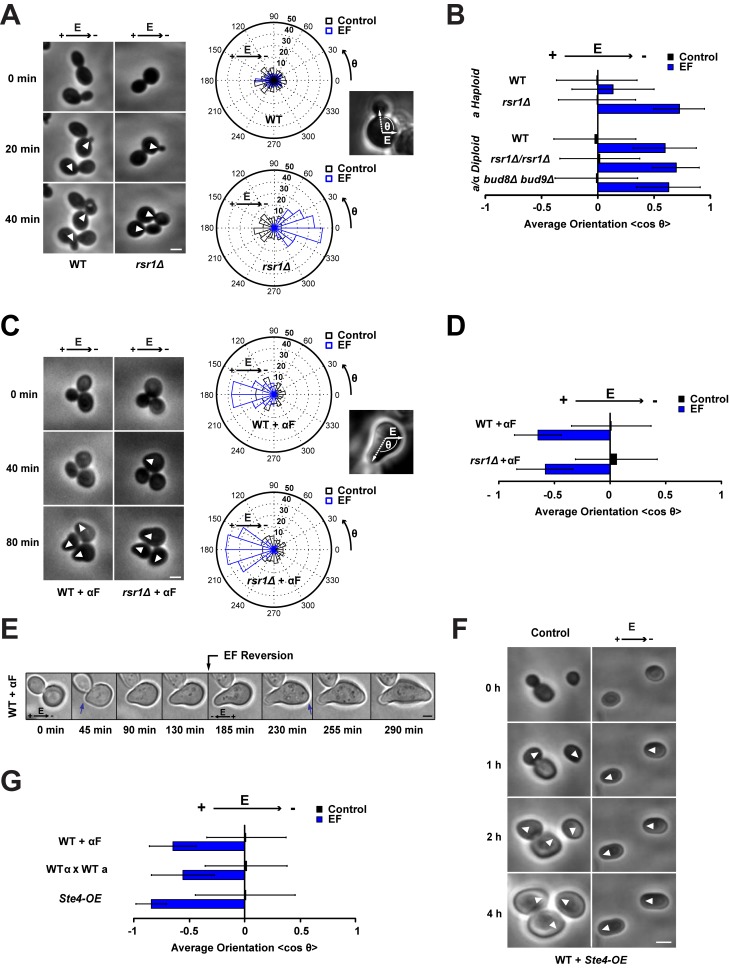
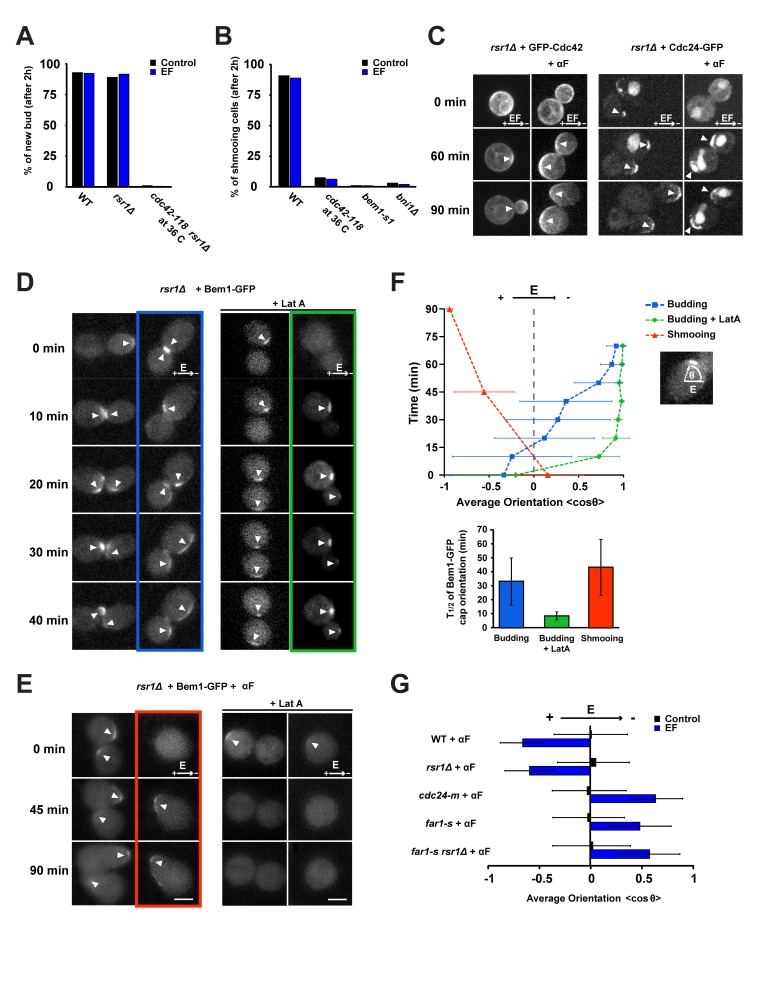
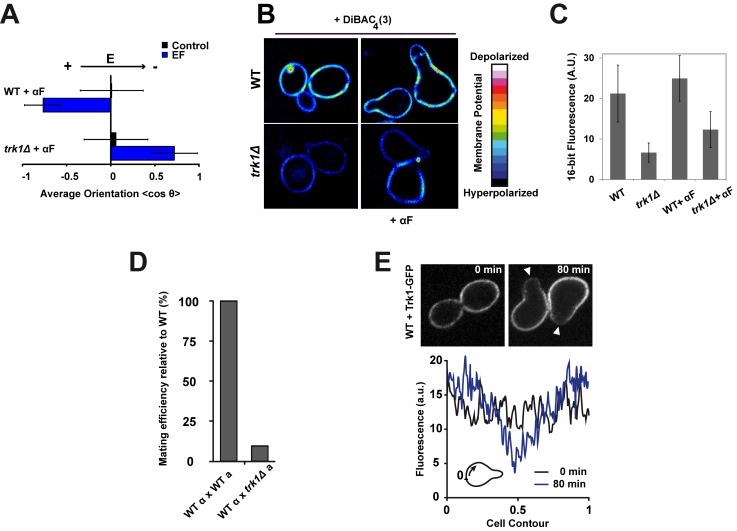
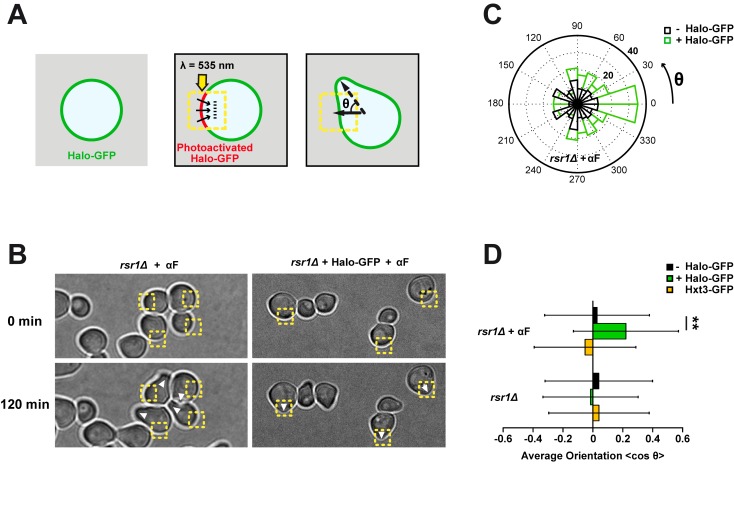
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
