

Soybean miR172c targets the repressive AP2 transcription factor NNC1 to activate ENOD40 expression and regulate nodule initiation
- PMID: 25549672
- PMCID: PMC4311200
- DOI: 10.1105/tpc.114.131607
Soybean miR172c targets the repressive AP2 transcription factor NNC1 to activate ENOD40 expression and regulate nodule initiation
Abstract
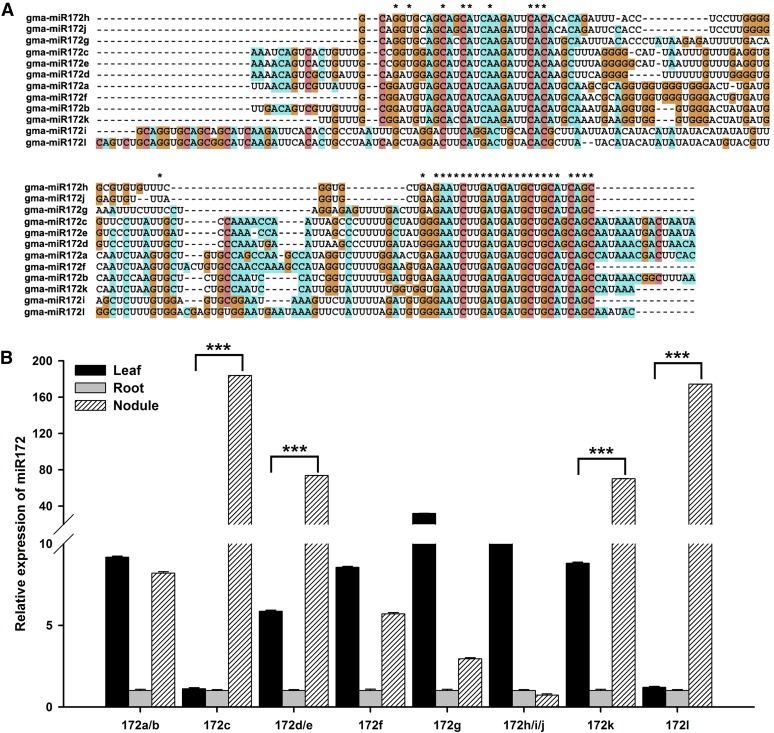
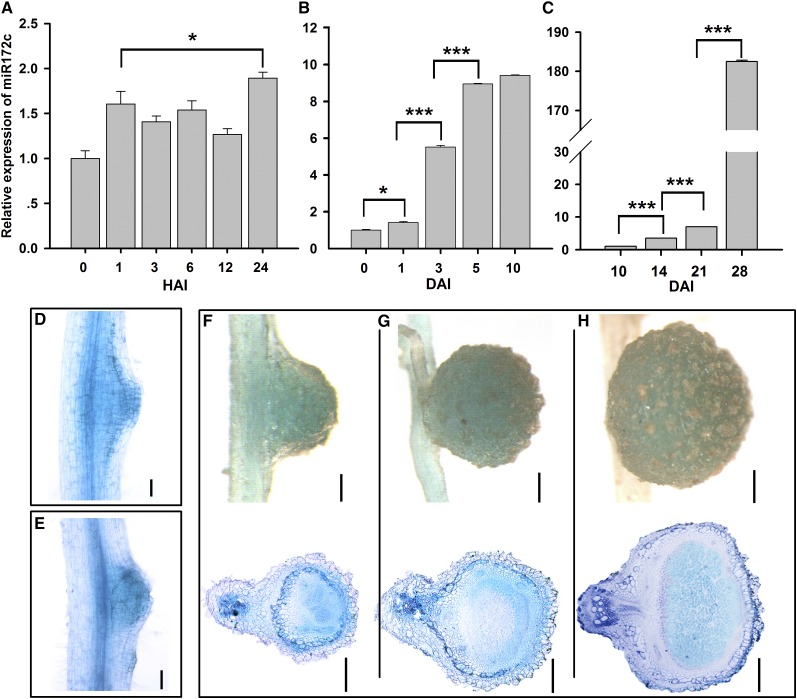
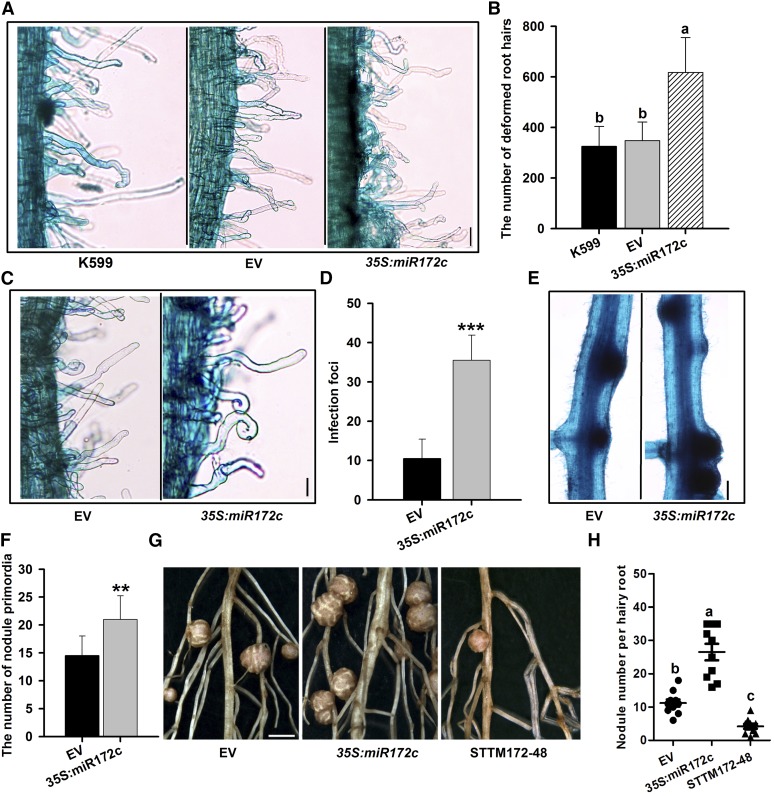
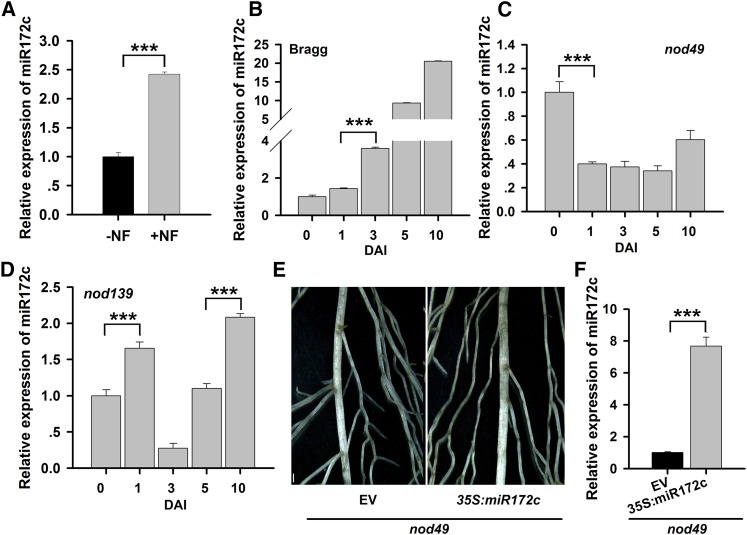
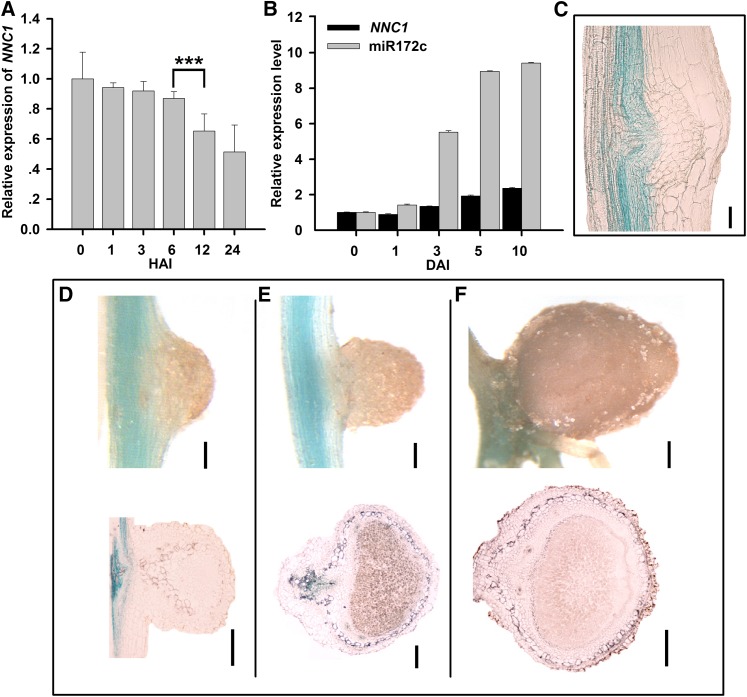
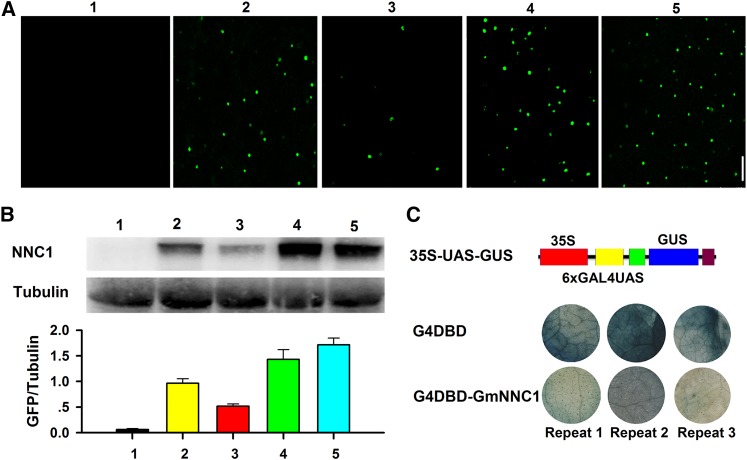
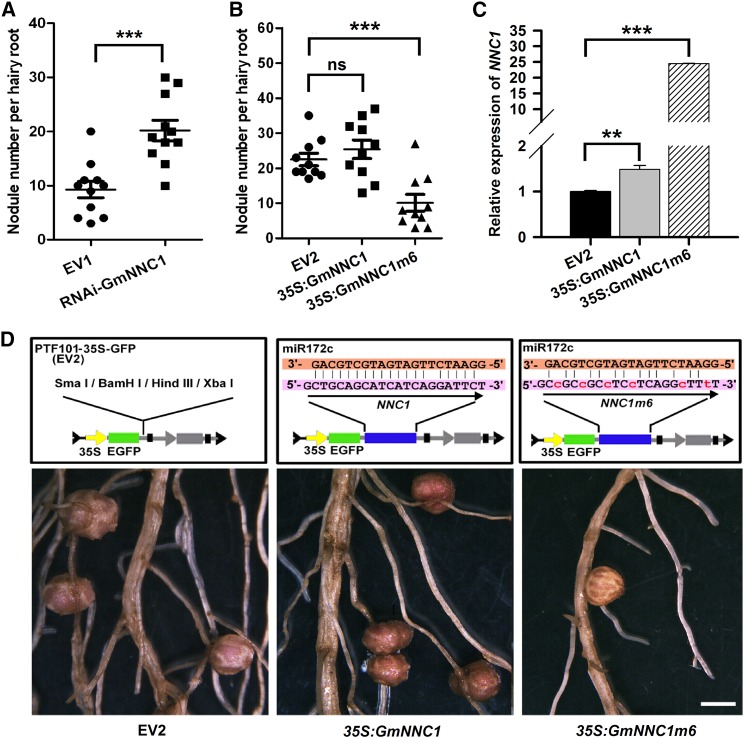
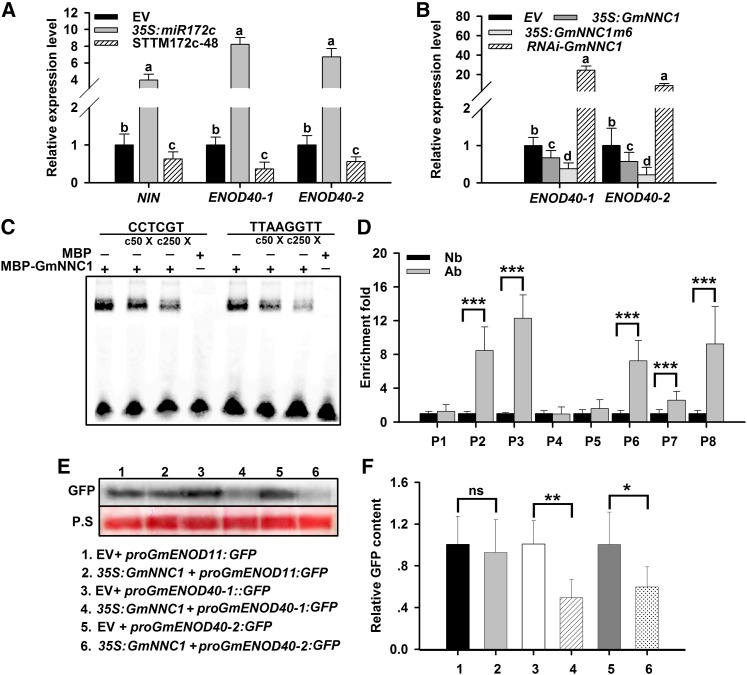
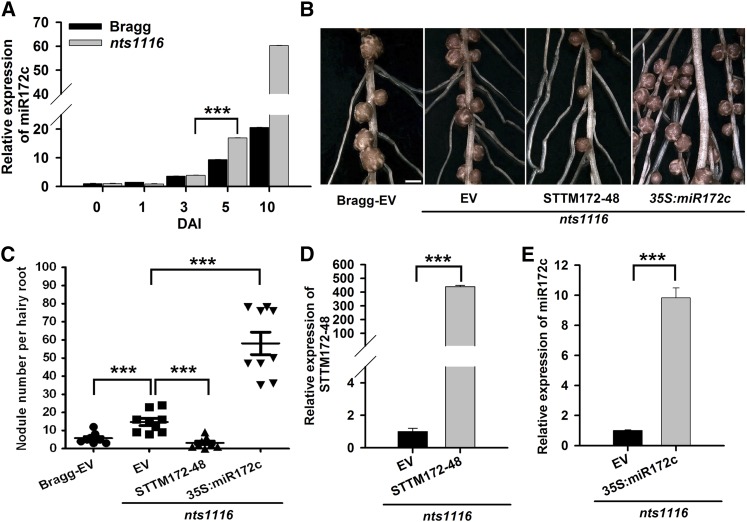
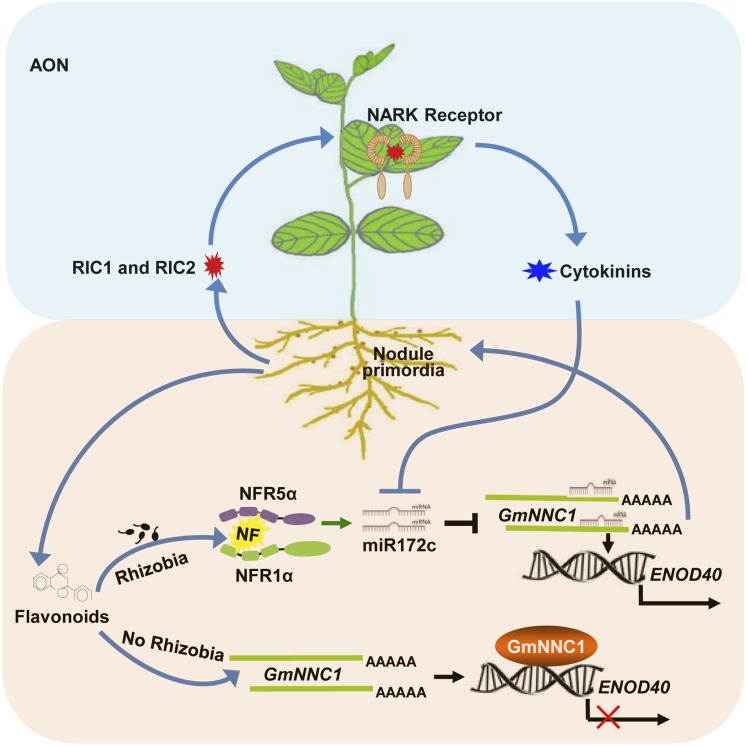
MicroRNAs are noncoding RNAs that act as master regulators to modulate various biological processes by posttranscriptionally repressing their target genes. Repression of their target mRNA(s) can modulate signaling cascades and subsequent cellular events. Recently, a role for miR172 in soybean (Glycine max) nodulation has been described; however, the molecular mechanism through which miR172 acts to regulate nodulation has yet to be explored. Here, we demonstrate that soybean miR172c modulates both rhizobium infection and nodule organogenesis. miR172c was induced in soybean roots inoculated with either compatible Bradyrhizobium japonicum or lipooligosaccharide Nod factor and was highly upregulated during nodule development. Reduced activity and overexpression of miR172c caused dramatic changes in nodule initiation and nodule number. We show that soybean miR172c regulates nodule formation by repressing its target gene, Nodule Number Control1, which encodes a protein that directly targets the promoter of the early nodulin gene, ENOD40. Interestingly, transcriptional levels of miR172c were regulated by both Nod Factor Receptor1α/5α-mediated activation and by autoregulation of nodulation-mediated inhibition. Thus, we established a direct link between miR172c and the Nod factor signaling pathway in addition to adding a new layer to the precise nodulation regulation mechanism of soybean.
© 2014 American Society of Plant Biologists. All rights reserved.
Figures
References
-
- Banfalvi Z., Nieuwkoop A., Schell M., Besl L., Stacey G. (1988). Regulation of nod gene expression in Bradyrhizobium japonicum. Mol. Gen. Genet. 214: 420–424. - PubMed
-
- Bustos-Sanmamed P., Mao G., Deng Y., Elouet M., Khan G.A., Bazin J., Lelandais-Brière C. (2013). Overexpression of miR160 affects root growth and nitrogen-fixing nodule number in Medicago truncatula. Funct. Plant Biol. 40: 1208–1220. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
