

The chemistry of negotiation: rhythmic, glycan-driven acidification in a symbiotic conversation
- PMID: 25550509
- PMCID: PMC4299225
- DOI: 10.1073/pnas.1418580112
The chemistry of negotiation: rhythmic, glycan-driven acidification in a symbiotic conversation
Abstract
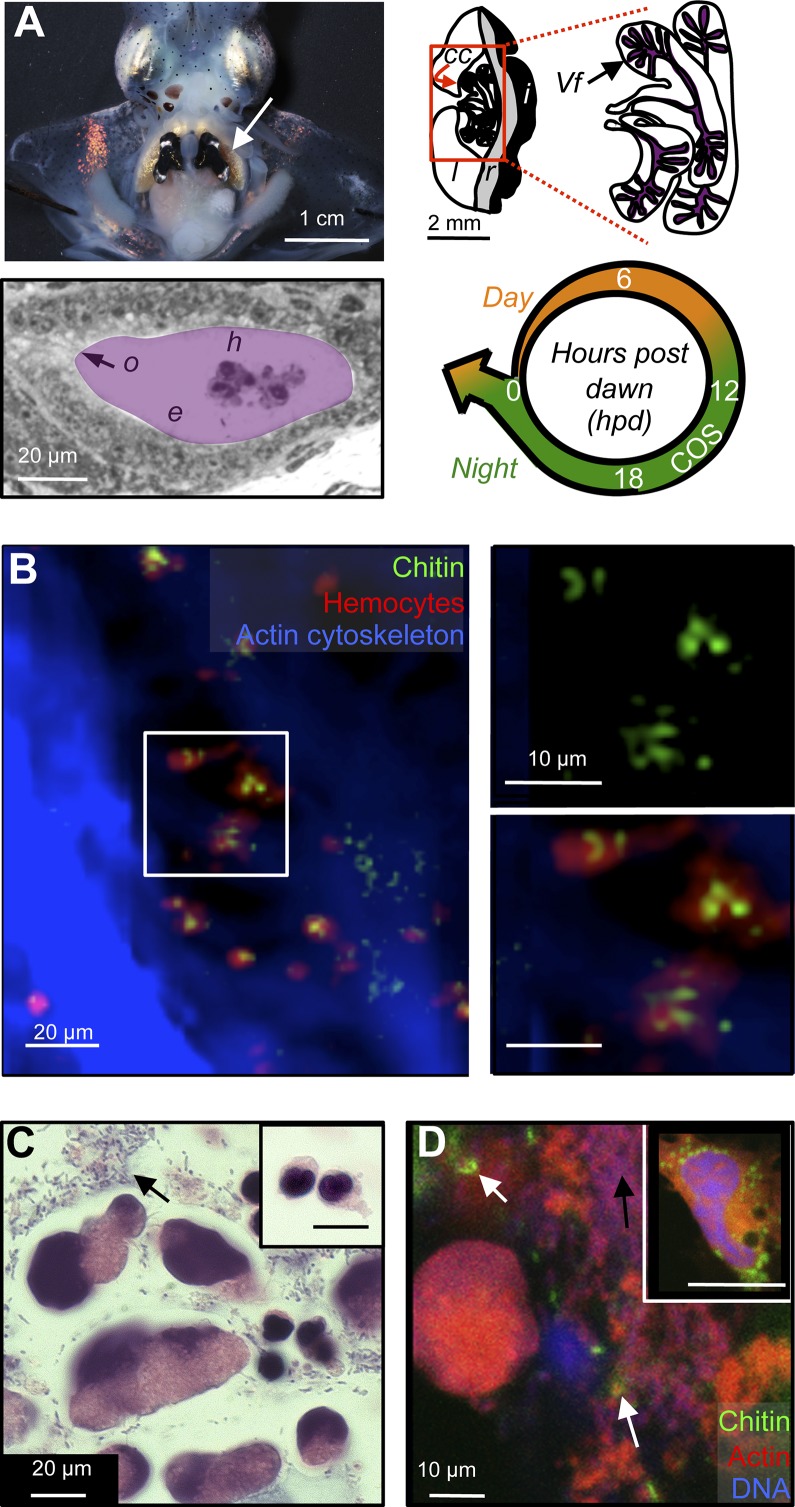
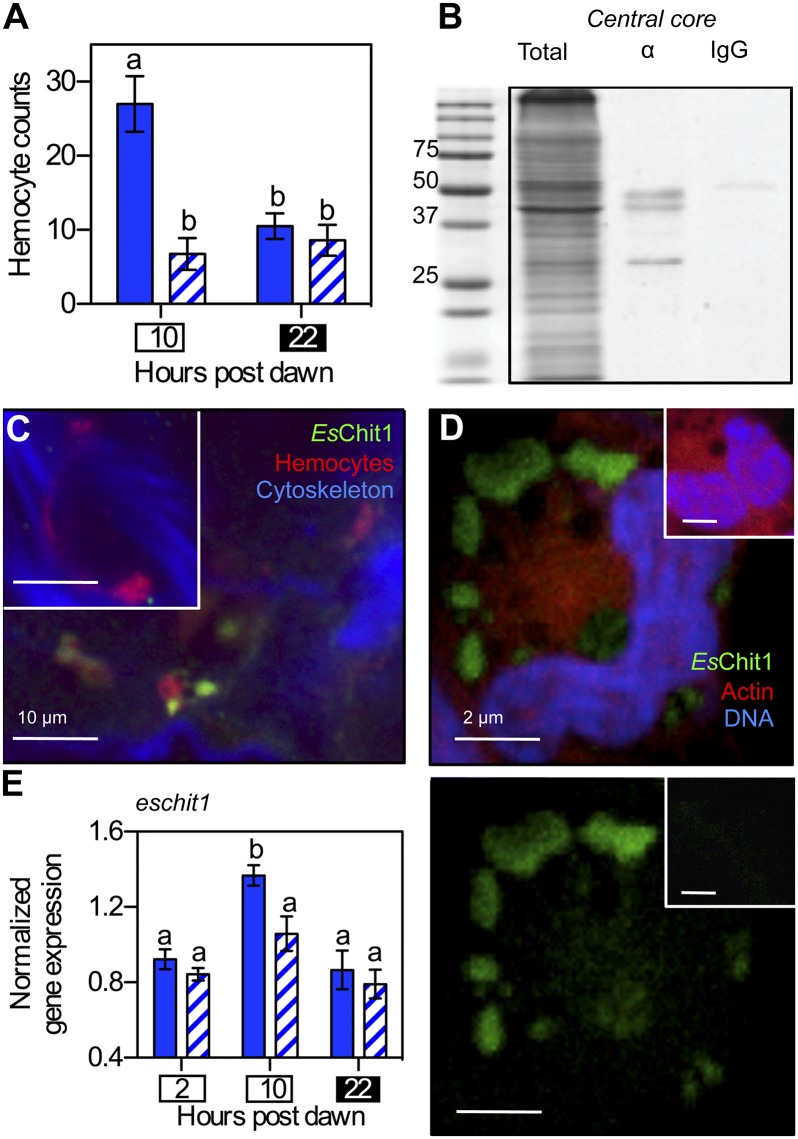
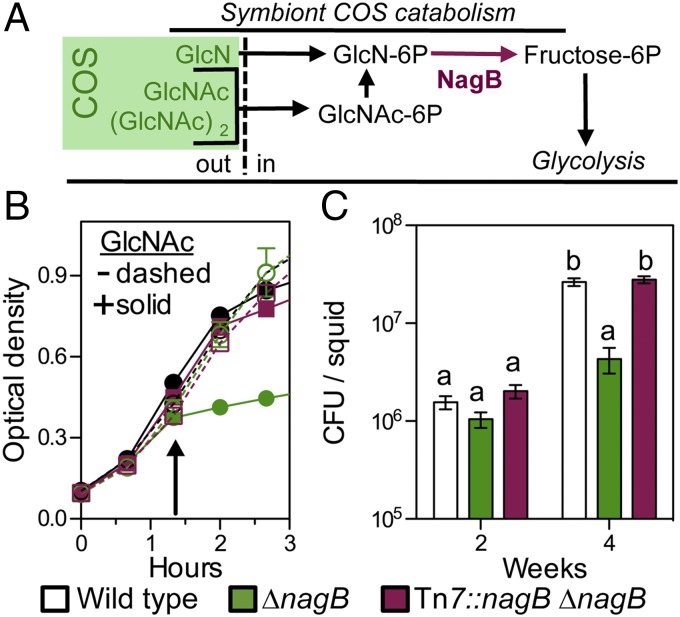
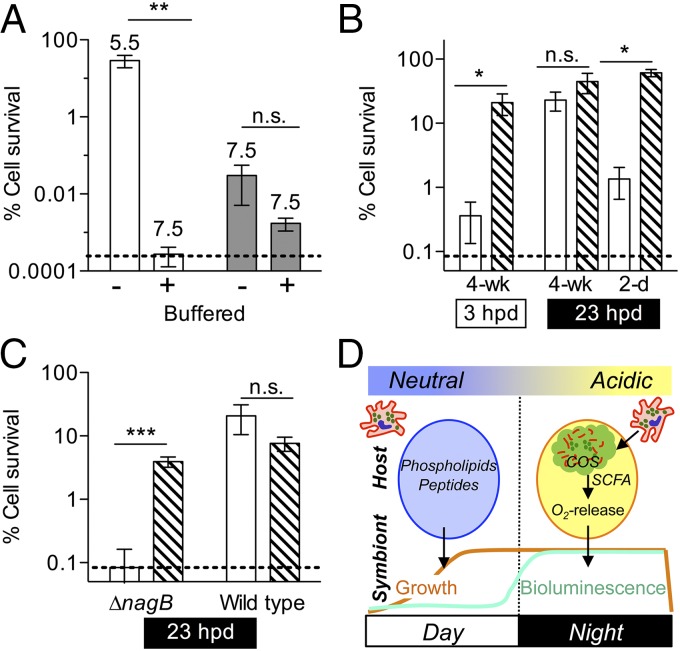
Glycans have emerged as critical determinants of immune maturation, microbial nutrition, and host health in diverse symbioses. In this study, we asked how cyclic delivery of a single host-derived glycan contributes to the dynamic stability of the mutualism between the squid Euprymna scolopes and its specific, bioluminescent symbiont, Vibrio fischeri. V. fischeri colonizes the crypts of a host organ that is used for behavioral light production. E. scolopes synthesizes the polymeric glycan chitin in macrophage-like immune cells called hemocytes. We show here that, just before dusk, hemocytes migrate from the vasculature into the symbiotic crypts, where they lyse and release particulate chitin, a behavior that is established only in the mature symbiosis. Diel transcriptional rhythms in both partners further indicate that the chitin is provided and metabolized only at night. A V. fischeri mutant defective in chitin catabolism was able to maintain a normal symbiont population level, but only until the symbiotic organ reached maturity (∼ 4 wk after colonization); this result provided a direct link between chitin utilization and symbiont persistence. Finally, catabolism of chitin by the symbionts was also specifically required for a periodic acidification of the adult crypts each night. This acidification, which increases the level of oxygen available to the symbionts, enhances their capacity to produce bioluminescence at night. We propose that other animal hosts may similarly regulate the activities of epithelium-associated microbial communities through the strategic provision of specific nutrients, whose catabolism modulates conditions like pH or anoxia in their symbionts' habitat.
Keywords: chitin; metabolism; squid–vibrio; symbiosis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Symbiosis: Sweet talking your partner.Nat Rev Microbiol. 2015 Feb;13(2):66-7. doi: 10.1038/nrmicro3424. Epub 2015 Jan 12. Nat Rev Microbiol. 2015. PMID: 25578956 No abstract available.
References
-
- Kereszt A, Mergaert P, Maróti G, Kondorosi E. Innate immunity effectors and virulence factors in symbiosis. Curr Opin Microbiol. 2011;14(1):76–81. - PubMed
-
- Axelrod R. The Evolution of Cooperation. Basic Books; New York: 1984.
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
