

Condensins exert force on chromatin-nuclear envelope tethers to mediate nucleoplasmic reticulum formation in Drosophila melanogaster
- PMID: 25552604
- PMCID: PMC4349088
- DOI: 10.1534/g3.114.015685
Condensins exert force on chromatin-nuclear envelope tethers to mediate nucleoplasmic reticulum formation in Drosophila melanogaster
Abstract
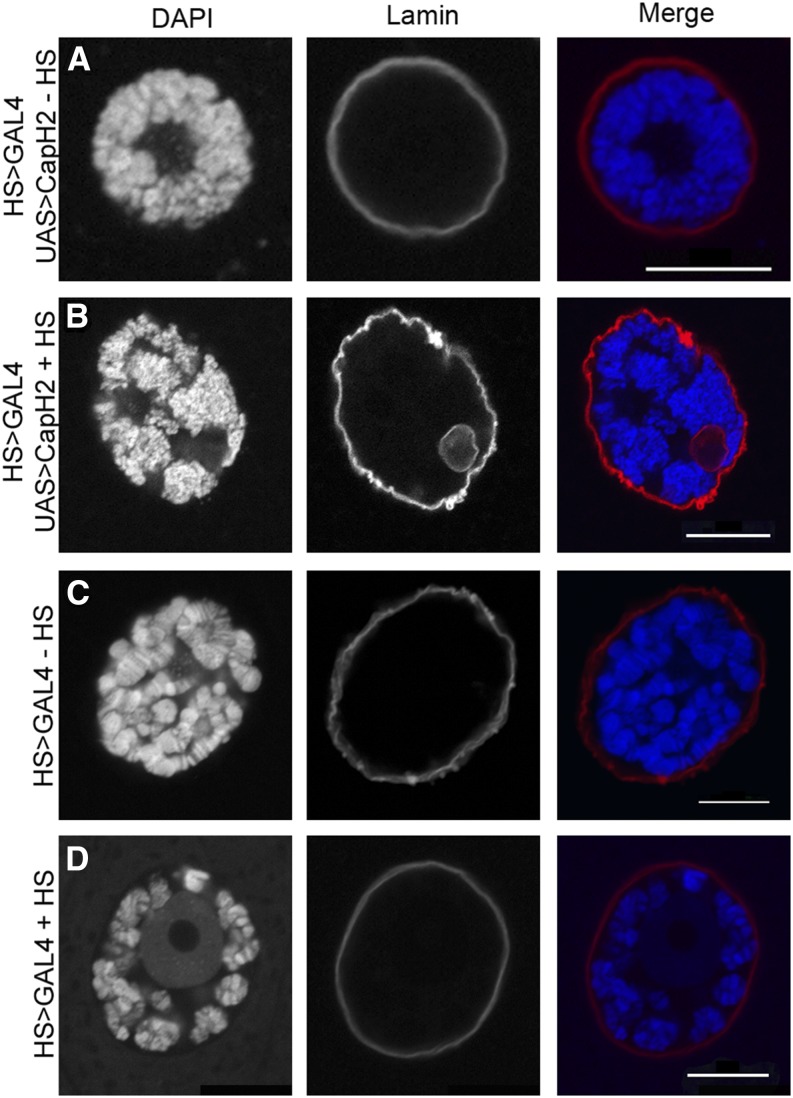
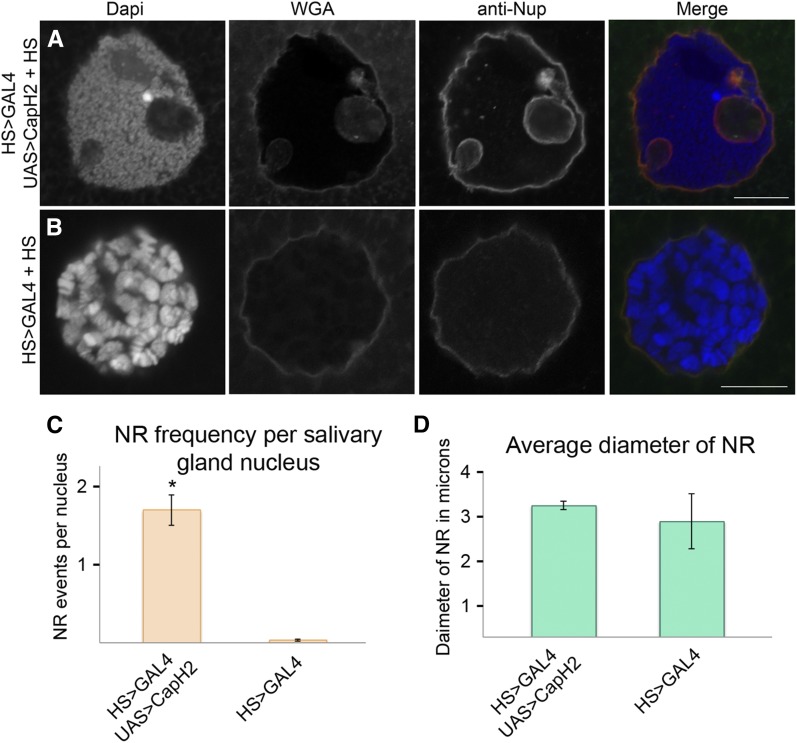
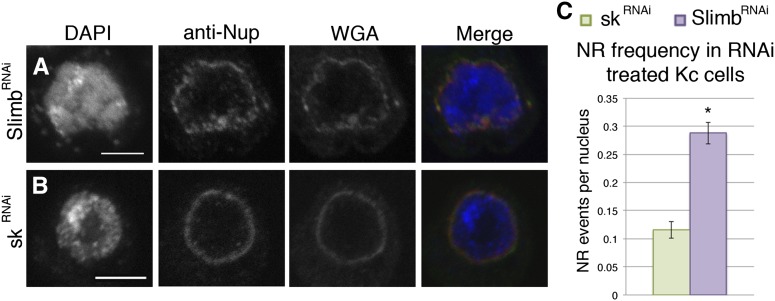
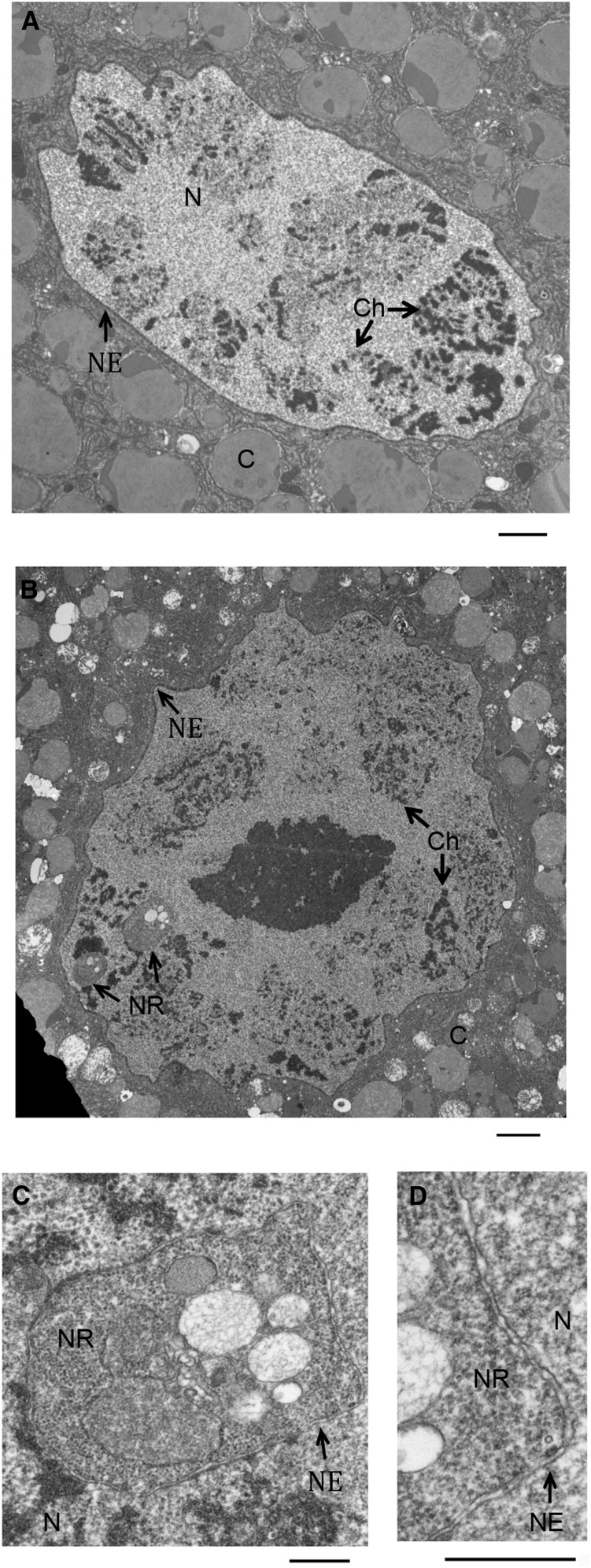
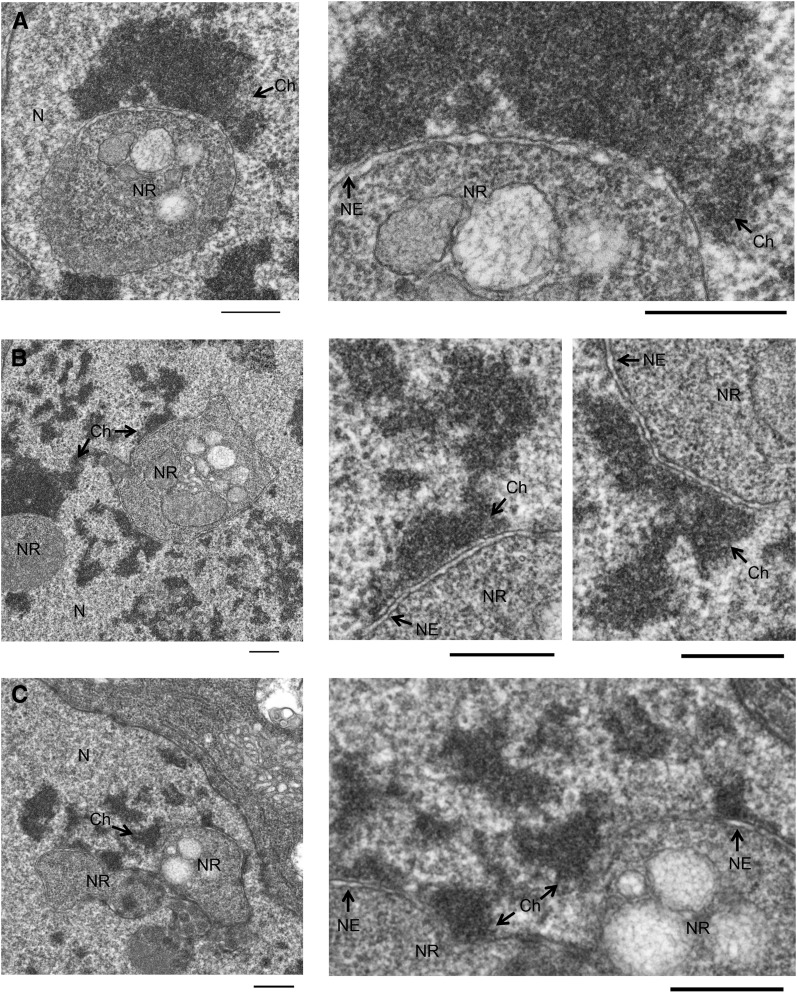
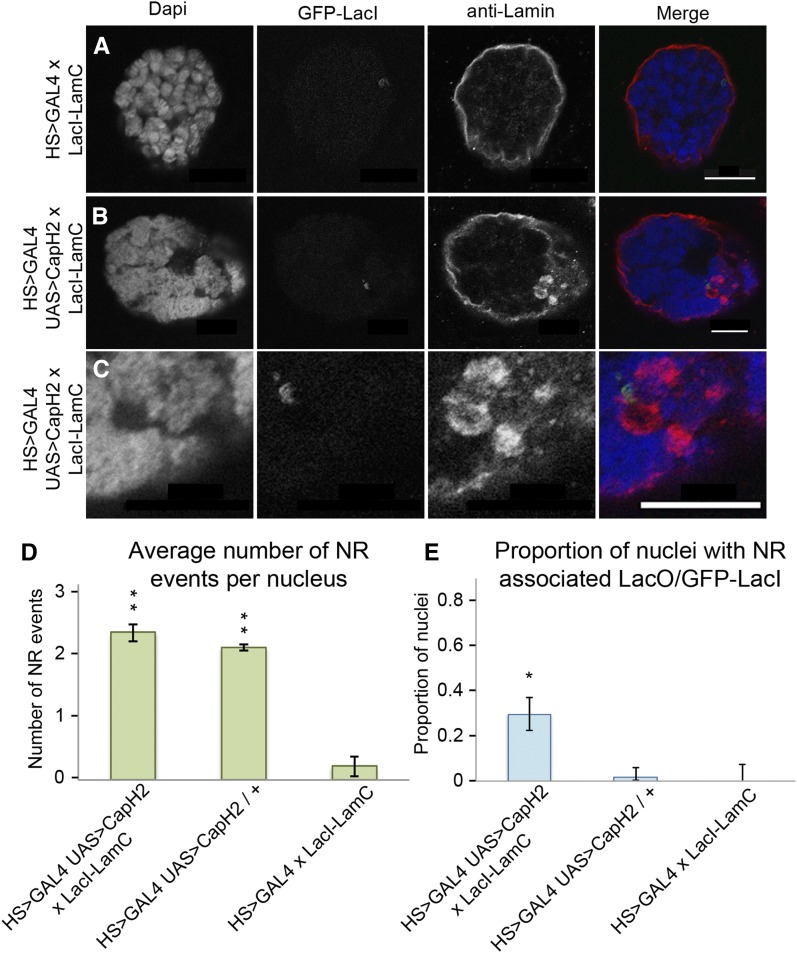
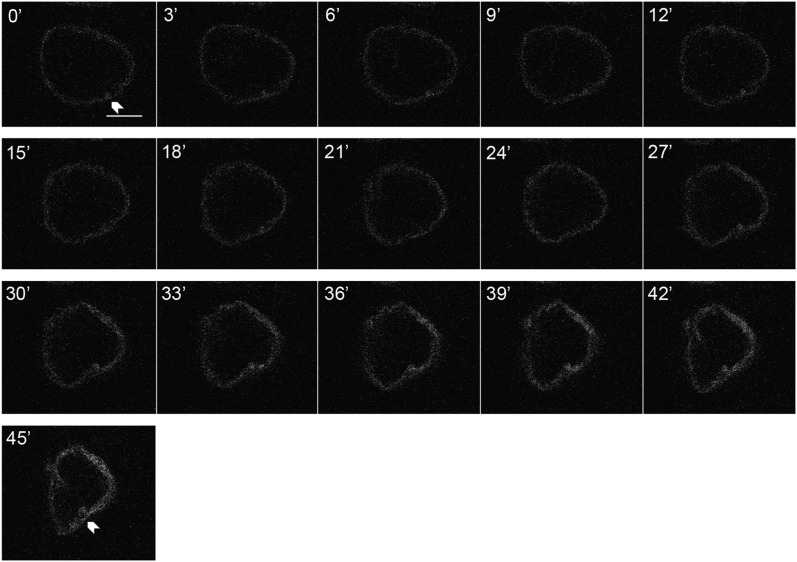
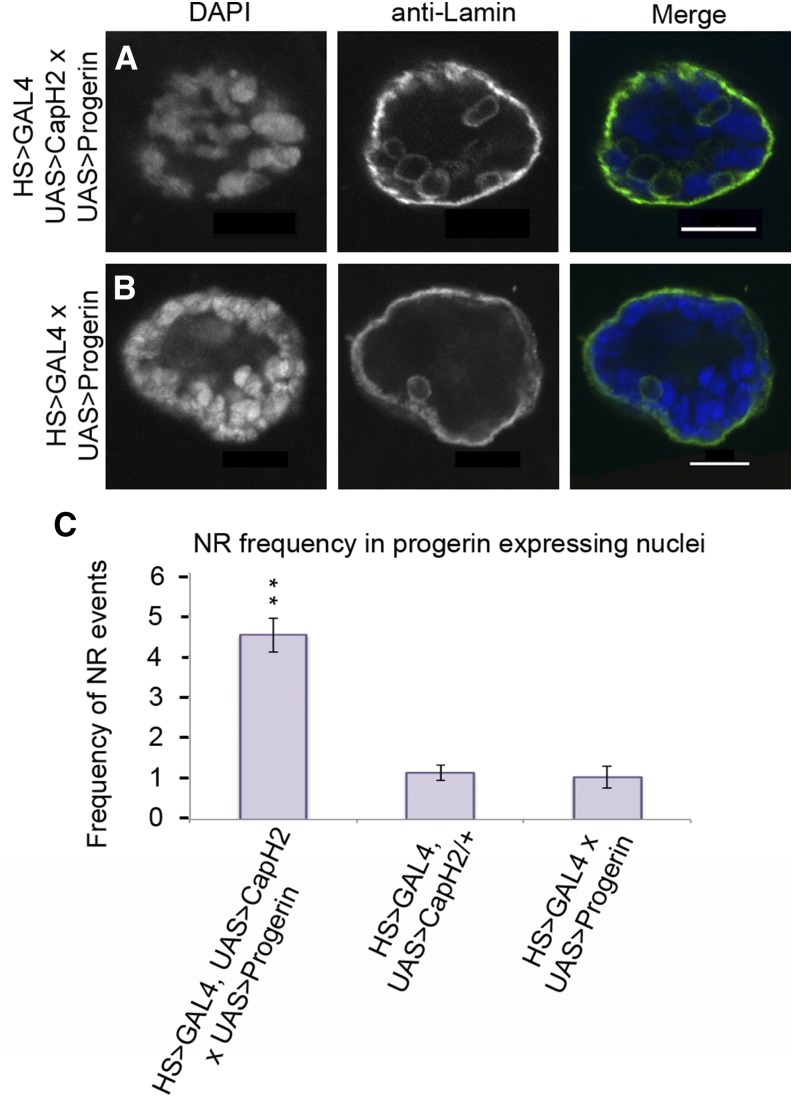
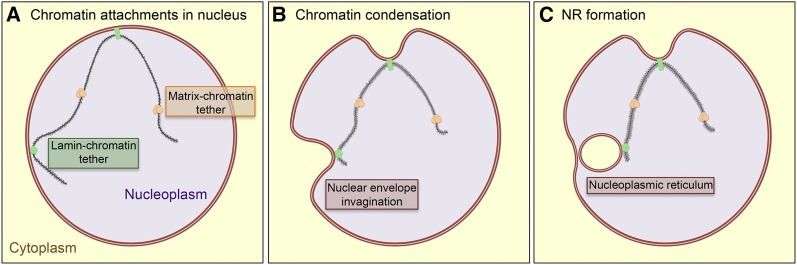
Although the nuclear envelope is known primarily for its role as a boundary between the nucleus and cytoplasm in eukaryotes, it plays a vital and dynamic role in many cellular processes. Studies of nuclear structure have revealed tissue-specific changes in nuclear envelope architecture, suggesting that its three-dimensional structure contributes to its functionality. Despite the importance of the nuclear envelope, the factors that regulate and maintain nuclear envelope shape remain largely unexplored. The nuclear envelope makes extensive and dynamic interactions with the underlying chromatin. Given this inexorable link between chromatin and the nuclear envelope, it is possible that local and global chromatin organization reciprocally impact nuclear envelope form and function. In this study, we use Drosophila salivary glands to show that the three-dimensional structure of the nuclear envelope can be altered with condensin II-mediated chromatin condensation. Both naturally occurring and engineered chromatin-envelope interactions are sufficient to allow chromatin compaction forces to drive distortions of the nuclear envelope. Weakening of the nuclear lamina further enhanced envelope remodeling, suggesting that envelope structure is capable of counterbalancing chromatin compaction forces. Our experiments reveal that the nucleoplasmic reticulum is born of the nuclear envelope and remains dynamic in that they can be reabsorbed into the nuclear envelope. We propose a model where inner nuclear envelope-chromatin tethers allow interphase chromosome movements to change nuclear envelope morphology. Therefore, interphase chromatin compaction may be a normal mechanism that reorganizes nuclear architecture, while under pathological conditions, such as laminopathies, compaction forces may contribute to defects in nuclear morphology.
Keywords: chromatin compaction; chromatin force; nuclear architecture; nuclear envelope; nucleus.
Copyright © 2015 Bozler et al.
Figures
Similar articles
-
Mechanisms for assembly of the nucleoplasmic reticulum.Cell Mol Life Sci. 2024 Oct 5;81(1):415. doi: 10.1007/s00018-024-05437-3. Cell Mol Life Sci. 2024. PMID: 39367888 Free PMC article. Review.
-
SCFSlimb ubiquitin ligase suppresses condensin II-mediated nuclear reorganization by degrading Cap-H2.J Cell Biol. 2013 Apr 1;201(1):49-63. doi: 10.1083/jcb.201207183. Epub 2013 Mar 25. J Cell Biol. 2013. PMID: 23530065 Free PMC article.
-
Maintenance of interphase chromosome compaction and homolog pairing in Drosophila is regulated by the condensin cap-h2 and its partner Mrg15.Genetics. 2013 Sep;195(1):127-46. doi: 10.1534/genetics.113.153544. Epub 2013 Jul 2. Genetics. 2013. PMID: 23821596 Free PMC article.
-
Condensin II Regulates Interphase Chromatin Organization Through the Mrg-Binding Motif of Cap-H2.G3 (Bethesda). 2015 Mar 9;5(5):803-17. doi: 10.1534/g3.115.016634. G3 (Bethesda). 2015. PMID: 25758823 Free PMC article.
-
Condensins and cohesins - one of these things is not like the other!J Cell Sci. 2019 Feb 7;132(3):jcs220491. doi: 10.1242/jcs.220491. J Cell Sci. 2019. PMID: 30733374 Free PMC article. Review.
Cited by
-
ATR-mediated regulation of nuclear and cellular plasticity.DNA Repair (Amst). 2016 Aug;44:143-150. doi: 10.1016/j.dnarep.2016.05.020. Epub 2016 May 16. DNA Repair (Amst). 2016. PMID: 27283761 Free PMC article. Review.
-
Mechanisms for assembly of the nucleoplasmic reticulum.Cell Mol Life Sci. 2024 Oct 5;81(1):415. doi: 10.1007/s00018-024-05437-3. Cell Mol Life Sci. 2024. PMID: 39367888 Free PMC article. Review.
-
Intranuclear inclusions in hepatocellular carcinoma contain autophagy-associated proteins and correlate with prolonged survival.J Pathol Clin Res. 2019 Jul;5(3):164-176. doi: 10.1002/cjp2.129. Epub 2019 Apr 8. J Pathol Clin Res. 2019. PMID: 30859721 Free PMC article.
-
Centripetal nuclear shape fluctuations associate with chromatin condensation in early prophase.Commun Biol. 2023 Jul 12;6(1):715. doi: 10.1038/s42003-023-05074-9. Commun Biol. 2023. PMID: 37438411 Free PMC article.
-
Heterochromatin, histone modifications, and nuclear architecture in disease vectors.Curr Opin Insect Sci. 2015 Aug 1;10:110-117. doi: 10.1016/j.cois.2015.05.003. Curr Opin Insect Sci. 2015. PMID: 26097808 Free PMC article.
References
-
- Beard G. S., Bridger J. M., Kill I. R., Tree D. R. P., 2008. Towards a Drosophila model of Hutchinson-Gilford progeria syndrome. Biochem. Soc. Trans. 36: 1389–1392. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases