

Potentially pathogenic immune cells and networks in apparently healthy lacrimal glands
- PMID: 25557346
- PMCID: PMC4284438
- DOI: 10.1016/j.jtos.2014.06.003
Potentially pathogenic immune cells and networks in apparently healthy lacrimal glands
Abstract
Lacrimal glands of people over 40 years old frequently contain lymphocytic infiltrates. Relationships between histopathological presentation and physiological dysfunction are not straightforward. Data from rabbit studies have suggested that at least two immune cell networks form in healthy lacrimal glands, one responding to environmental dryness, the other to high temperatures. New findings indicate that mRNAs for several chemokines and cytokines are expressed primarily in epithelial cells; certain others are expressed in both epithelial cells and immune cells. Transcript abundances vary substantially across glands from animals that have experienced the same conditions, allowing for correlation analyses, which detect clusters that map to various cell types and to networks of coordinately functioning cells. A core network--expressing mRNAs including IL-1α, IL-6, IL-17A, and IL-10--expands adaptively with exposure to dryness, suppressing IFN-γ, but potentially causing physiological dysfunction. High temperature elicits concurrent increases of mRNAs for prolactin (PRL), CCL21, and IL-18. PRL is associated with crosstalk to IFN-γ, BAFF, and IL-4. The core network reacts to the resulting PRL-BAFF-IL-4 network, creating a profile reminiscent of Sjögren's disease. In a warmer, moderately dry setting, PRL-associated increases of IFN-γ are associated with suppression of IL-10 and augmentations of IL-1α and IL-17, creating a profile reminiscent of severe chronic inflammation.
Keywords: Sjögren's disease(∗); aging; autoimmunity; chronic inflammation; dacryoadenitis; dry eye; prolactin.
Copyright © 2015 The Authors. Published by Elsevier Inc. All rights reserved.
Figures







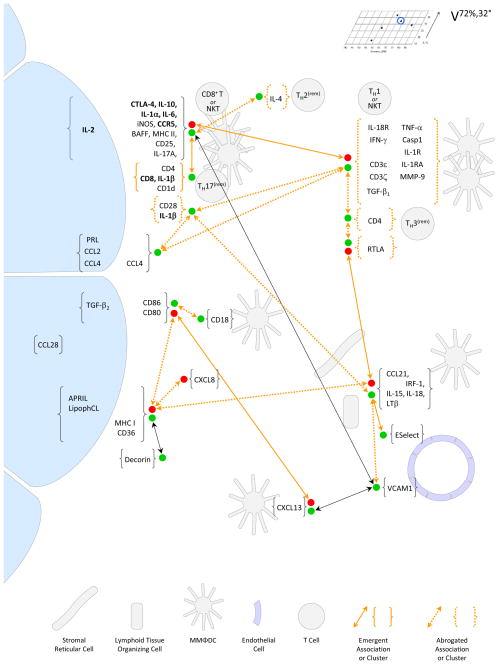

 ) was below the limit of detection. Details are as described in the legend to Figure 4. Development of the core, reactive-adaptive network was associated with positive crosstalk to TH2-reminiscent cells, which expressed IL-4; with positive crosstalk to cells that expressed CCL21; and, indirectly, with positive crosstalk to cells that expressed CXCL13. As shown in Figure 11, none of the glands presented with frank immunopathology, but as discussed in Section III, increasing levels of iNOS, IL-1, and IL-6 may come to be associated with physiological dysfunction.
) was below the limit of detection. Details are as described in the legend to Figure 4. Development of the core, reactive-adaptive network was associated with positive crosstalk to TH2-reminiscent cells, which expressed IL-4; with positive crosstalk to cells that expressed CCL21; and, indirectly, with positive crosstalk to cells that expressed CXCL13. As shown in Figure 11, none of the glands presented with frank immunopathology, but as discussed in Section III, increasing levels of iNOS, IL-1, and IL-6 may come to be associated with physiological dysfunction.

 ) were omitted from the initial regression analyses to minimize both type 1 and type 2 errors. Solid lines indicate range of X-axis values over which regressions were calculated, and dashed lines indicate projections. A. Associations with CD4 mRNA. The number of T cells (RTLA+ cells) and the abundance of CCL21 mRNA increased in approximately constant proportions across the entire sample of glands. Exponential regressions of IL-4 and MMP-9 mRNA abundances across the first five glands fell short of the P <.05 criterion for statistical significance. However, projections of both predicted values in gland V61%,27°01.OS that were near the observed values, and regressions calculated across all six glands were highly significant (dotted lines). B. Lack of a significant association between the abundances of mRNAs for CD4 and IL-10 across the first five V61%,27° glands. C. Associations with IL-10 mRNA. With the exception of CCR5 mRNA, the abundances of core, adaptive-reactive network transcripts in gland V61%,27°02.OS fell within two standard errors of the proportions in the other V61%,27° glands. This finding suggests that the network remained intact as it reacted to positive crosstalk associated with the immunopathological process. Abundances of mRNAs for CCL4, CCL2, CD25, CD28, BAFF, and numbers of bone marrow-derived cells—marked by CD18—were not significantly associated with the core, adaptive-reactive network transcripts across the first five glands but were significantly increased in gland V61%,27°02.OS.
) were omitted from the initial regression analyses to minimize both type 1 and type 2 errors. Solid lines indicate range of X-axis values over which regressions were calculated, and dashed lines indicate projections. A. Associations with CD4 mRNA. The number of T cells (RTLA+ cells) and the abundance of CCL21 mRNA increased in approximately constant proportions across the entire sample of glands. Exponential regressions of IL-4 and MMP-9 mRNA abundances across the first five glands fell short of the P <.05 criterion for statistical significance. However, projections of both predicted values in gland V61%,27°01.OS that were near the observed values, and regressions calculated across all six glands were highly significant (dotted lines). B. Lack of a significant association between the abundances of mRNAs for CD4 and IL-10 across the first five V61%,27° glands. C. Associations with IL-10 mRNA. With the exception of CCR5 mRNA, the abundances of core, adaptive-reactive network transcripts in gland V61%,27°02.OS fell within two standard errors of the proportions in the other V61%,27° glands. This finding suggests that the network remained intact as it reacted to positive crosstalk associated with the immunopathological process. Abundances of mRNAs for CCL4, CCL2, CD25, CD28, BAFF, and numbers of bone marrow-derived cells—marked by CD18—were not significantly associated with the core, adaptive-reactive network transcripts across the first five glands but were significantly increased in gland V61%,27°02.OS.






Similar articles
-
Molecular Evidence for Precursors of Sjögren's Foci in Histologically Normal Lacrimal Glands.Int J Mol Sci. 2019 Jan 8;20(1):223. doi: 10.3390/ijms20010223. Int J Mol Sci. 2019. PMID: 30626116 Free PMC article.
-
Matriptase deletion initiates a Sjögren's syndrome-like disease in mice.PLoS One. 2014 Feb 13;9(2):e82852. doi: 10.1371/journal.pone.0082852. eCollection 2014. PLoS One. 2014. PMID: 24551030 Free PMC article.
-
Interferon-γ treatment in vitro elicits some of the changes in cathepsin S and antigen presentation characteristic of lacrimal glands and corneas from the NOD mouse model of Sjögren's Syndrome.PLoS One. 2017 Sep 13;12(9):e0184781. doi: 10.1371/journal.pone.0184781. eCollection 2017. PLoS One. 2017. PMID: 28902875 Free PMC article.
-
Immunopathology of Sjögren's syndrome.Ann Med Interne (Paris). 1998 Feb;149(1):17-24. Ann Med Interne (Paris). 1998. PMID: 11490512 Review.
-
The geoepidemiology of Sjögren's syndrome.Autoimmun Rev. 2010 Mar;9(5):A305-10. doi: 10.1016/j.autrev.2009.11.004. Epub 2009 Nov 10. Autoimmun Rev. 2010. PMID: 19903539 Review.
Cited by
-
Molecular Evidence for Precursors of Sjögren's Foci in Histologically Normal Lacrimal Glands.Int J Mol Sci. 2019 Jan 8;20(1):223. doi: 10.3390/ijms20010223. Int J Mol Sci. 2019. PMID: 30626116 Free PMC article.
-
High-Fat Nutritional Challenge Reshapes Circadian Signatures in Murine Extraorbital Lacrimal Glands.Invest Ophthalmol Vis Sci. 2022 May 2;63(5):23. doi: 10.1167/iovs.63.5.23. Invest Ophthalmol Vis Sci. 2022. PMID: 35588356 Free PMC article.
-
Multiple Natural and Experimental Inflammatory Rabbit Lacrimal Gland Phenotypes.Ocul Surf. 2016 Oct;14(4):460-483.e3. doi: 10.1016/j.jtos.2016.07.001. Epub 2016 Jul 15. Ocul Surf. 2016. PMID: 27423911 Free PMC article.
-
Bibliometric and visualized analysis of research hotpots and future prospects in lacrimal duct obstruction disease (1900 to 2024).Int J Ophthalmol. 2025 Mar 18;18(3):526-540. doi: 10.18240/ijo.2025.03.21. eCollection 2025. Int J Ophthalmol. 2025. PMID: 40103946 Free PMC article.
References
-
- Sjögren H, Bloch KJ. Keratoconjunctivitis sicca and Sjögren’s syndrome. Surv Ophthalmol. 1971;16:145–59.
-
- Sato EA, Matsumoto Y, Dogru M, et al. Lacrimal gland in Sjögren’s syndrome. Ophthalmology. 2010;117:1055–1055. e3. - PubMed
-
- Ogawa Y, Kuwana M. Dry eye as a major complication associated with chronic graft-versus-host disease after hematopoietic stem cell transplantation. Cornea. 2003;22 (Suppl 1):S19–27. - PubMed
-
- Yanardag H, Pamuk ÖN. Lacrimal gland involvement in sarcoidosis. Swiss Med Wkly. 2003;133:388–91. - PubMed
-
- Hunter DG, Foster CS. Ocular manifestations of sarcoidosis. In: Albert DM, Jakobiec FA, editors. Principles and Practice of Ophthalmology. Philadelphia: WB Saunders; 1994. pp. 443–50.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
