

Hypersensitive response-like lesions 1 codes for AtPPT1 and regulates accumulation of ROS and defense against bacterial pathogen Pseudomonas syringae in Arabidopsis thaliana
- PMID: 25557512
- PMCID: PMC4361009
- DOI: 10.1089/ars.2014.5963
Hypersensitive response-like lesions 1 codes for AtPPT1 and regulates accumulation of ROS and defense against bacterial pathogen Pseudomonas syringae in Arabidopsis thaliana
Abstract
Aims: Plants employ both basal and resistance gene (R gene)-mediated defenses in response to pathogens. Reactive oxygen species (ROS) are widely reported to play a central role in both basal and R gene-mediated defense; however, the nature of ROS has been less well established for basal defense. In addition, spatial distribution of redox moieties and mechanisms of plant responses during basal defense are poorly understood. We investigated redox signaling in Arabidopsis thaliana in response to virulent bacterial pathogen, focusing on the role of the mitochondria in balancing energy demands against generation of physiologically relevant ROS.
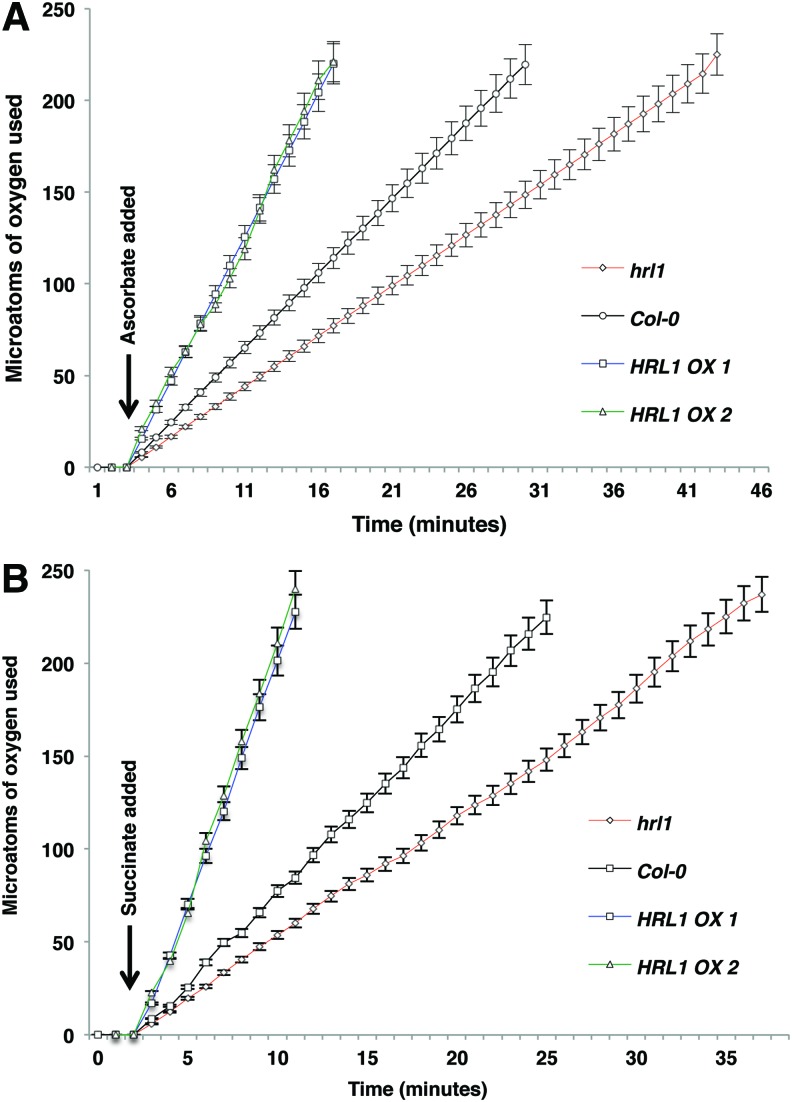
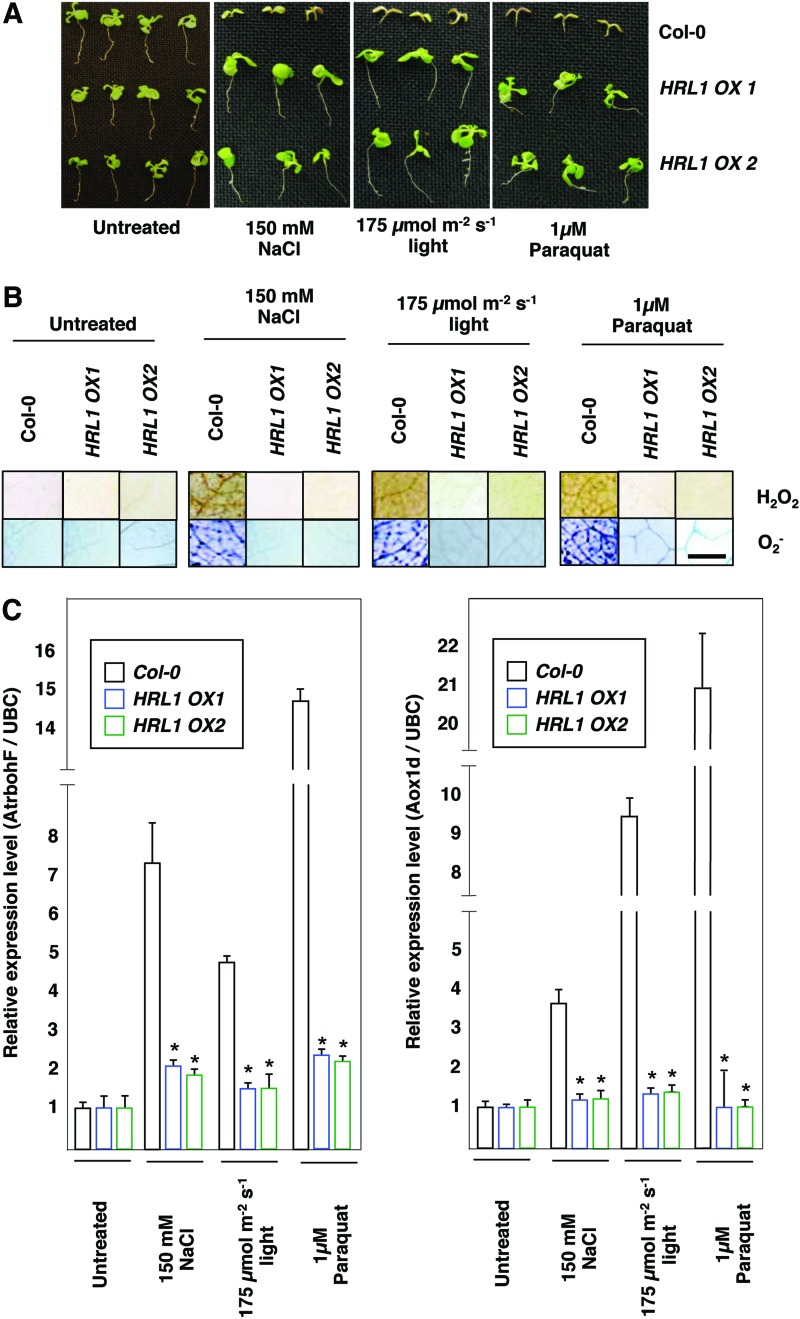
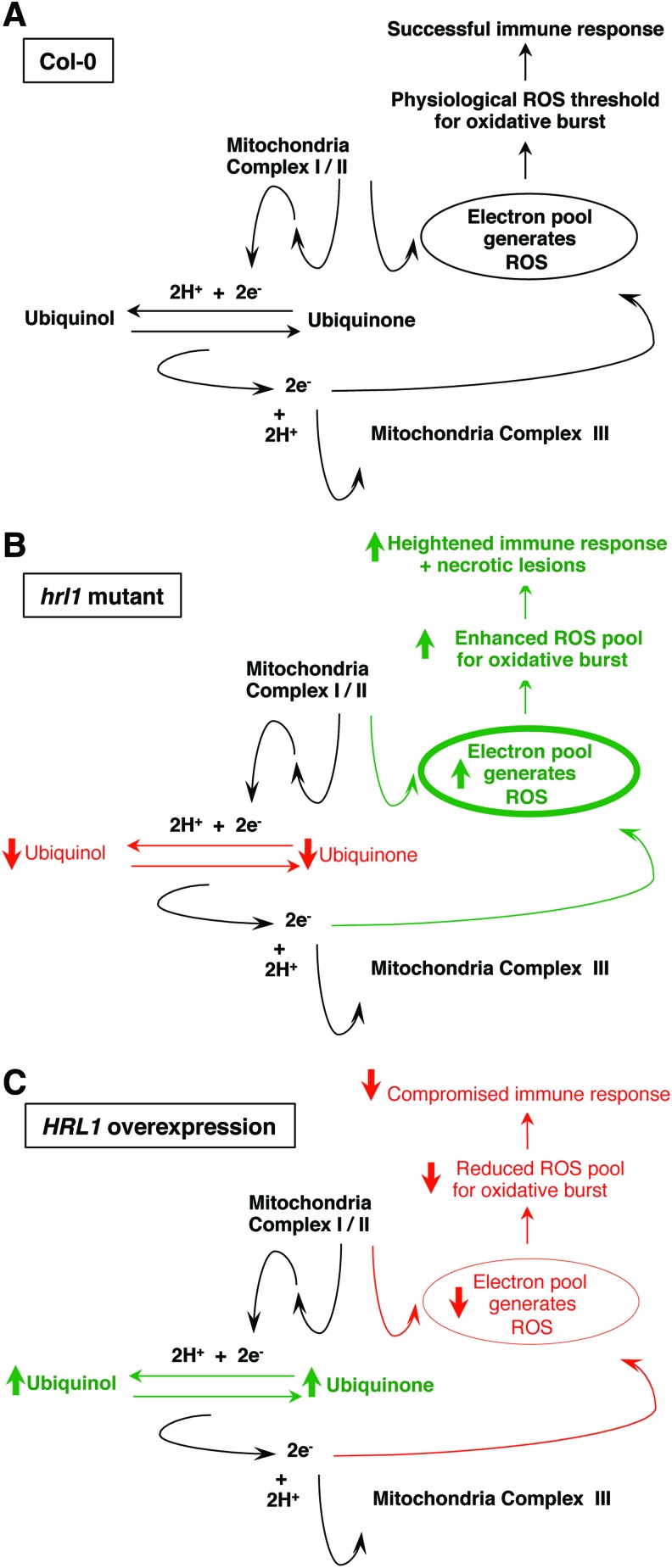
Results: Positional cloning of an Arabidopsis lesion mimic mutant identified a polyprenyl transferase involved in the biosynthesis of Coenzyme Q10 (CoQ), which leads to novel insights into physiological ROS levels and their role in basal resistance. Gain- and loss-of-function studies identified Coenzyme Q10 redox state to be a key determinant of ROS levels. These Coenzyme Q10 redox state-mediated ROS levels had a direct bearing on both response against pathogen and ability to thrive in high oxidative stress environments.
Innovation: We demonstrate that Coenzyme Q10 redox state generates an ROS threshold for a successful basal resistance response. Perturbation of the Coenzyme Q10 redox state has the potential to disrupt plant defense responses against bacterial pathogens.
Conclusions: Coenzyme Q10 redox state is a key regulator of Arabidopsis basal resistance against bacterial pathogens.
Figures
Similar articles
-
The secondary metabolism glycosyltransferases UGT73B3 and UGT73B5 are components of redox status in resistance of Arabidopsis to Pseudomonas syringae pv. tomato.Plant Cell Environ. 2014 May;37(5):1114-29. doi: 10.1111/pce.12221. Epub 2013 Nov 24. Plant Cell Environ. 2014. PMID: 24131360
-
AtRbohF is a crucial modulator of defence-associated metabolism and a key actor in the interplay between intracellular oxidative stress and pathogenesis responses in Arabidopsis.Plant J. 2012 Feb;69(4):613-27. doi: 10.1111/j.1365-313X.2011.04816.x. Epub 2011 Nov 25. Plant J. 2012. PMID: 21985584
-
The Arabidopsis ATAF1, a NAC transcription factor, is a negative regulator of defense responses against necrotrophic fungal and bacterial pathogens.Mol Plant Microbe Interact. 2009 Oct;22(10):1227-38. doi: 10.1094/MPMI-22-10-1227. Mol Plant Microbe Interact. 2009. PMID: 19737096
-
Role of proline and pyrroline-5-carboxylate metabolism in plant defense against invading pathogens.Front Plant Sci. 2015 Jul 6;6:503. doi: 10.3389/fpls.2015.00503. eCollection 2015. Front Plant Sci. 2015. PMID: 26217357 Free PMC article. Review.
-
Coenzyme Q redox signalling and longevity.Free Radic Biol Med. 2021 Feb 20;164:187-205. doi: 10.1016/j.freeradbiomed.2021.01.018. Epub 2021 Jan 12. Free Radic Biol Med. 2021. PMID: 33450379 Review.
Cited by
-
The Transcriptional Responses of Ectomycorrhizal Fungus, Cenococcum geophilum, to Drought Stress.J Fungi (Basel). 2022 Dec 21;9(1):15. doi: 10.3390/jof9010015. J Fungi (Basel). 2022. PMID: 36675836 Free PMC article.
-
Novel Insights into Floral Thermogenesis: In Vivo Analyses of Mitochondrial Dynamics in Nelumbo nucifera Flowers.Int J Mol Sci. 2022 Oct 8;23(19):11950. doi: 10.3390/ijms231911950. Int J Mol Sci. 2022. PMID: 36233249 Free PMC article.
-
SmPPT, a 4-hydroxybenzoate polyprenyl diphosphate transferase gene involved in ubiquinone biosynthesis, confers salt tolerance in Salvia miltiorrhiza.Plant Cell Rep. 2019 Dec;38(12):1527-1540. doi: 10.1007/s00299-019-02463-5. Epub 2019 Aug 30. Plant Cell Rep. 2019. PMID: 31471635
-
Plastoquinone and Ubiquinone in Plants: Biosynthesis, Physiological Function and Metabolic Engineering.Front Plant Sci. 2016 Dec 16;7:1898. doi: 10.3389/fpls.2016.01898. eCollection 2016. Front Plant Sci. 2016. PMID: 28018418 Free PMC article. Review.
-
Arabidopsis SMALL DEFENSE-ASSOCIATED PROTEIN 1 Modulates Pathogen Defense and Tolerance to Oxidative Stress.Front Plant Sci. 2020 Jun 3;11:703. doi: 10.3389/fpls.2020.00703. eCollection 2020. Front Plant Sci. 2020. PMID: 32582244 Free PMC article.
References
-
- Belkhadir Y, Subramaniam R, and Dangl JL. Plant disease resistance protein signaling: NBS-LRR proteins and their partners. Curr Opin Plant Biol 7: 391–399, 2004 - PubMed
-
- Bradley DJ, Kjellbom P, and Lamb CJ. Elicitor- and wound-induced oxidative cross-linking of a proline-rich plant cell wall protein: a novel, rapid defense response. Cell 70: 21–30, 1992 - PubMed
-
- Braidot E, Petrussa E, Vianello A, and Macri F. Hydrogen peroxide generation by higher plant mitochondria oxidizing complex I or complex II substrates. FEBS Lett 451: 347–350, 1999 - PubMed
-
- Chance B. and Williams GR. The respiratory chain and oxidative phosphorylation. Adv Enzymol Relat Subj Biochem 17: 65–134, 1956 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
