

Review
doi: 10.1152/physiol.00033.2014.
Aging and the muscle-bone relationship
Affiliations
- PMID: 25559151
- PMCID: PMC4285576
- DOI: 10.1152/physiol.00033.2014
Item in Clipboard
Review
Aging and the muscle-bone relationship
Physiology (Bethesda).
2015 Jan.
Abstract
Aging-induced declines in muscle size and quality are thought to contribute to catabolic alterations in bone, but changes in bone with age also profoundly alter its response to muscle-derived stimuli. This review provides an overview of some of the alterations that occur in muscle and bone with aging, and discusses the cellular and molecular mechanisms that may impact these age-associated changes.
©2015 Int. Union Physiol. Sci./Am. Physiol. Soc.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the author(s).
Figures
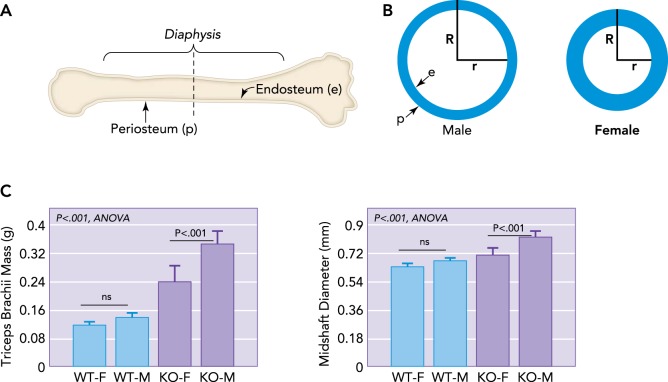
Schematic representation of a long bone and dimorphism in bone cross-sectional geometry A: schematic representation of a long bone indicating the outer periosteal surface (p) and inner endocortical surface lining the medullary cavity (e), with a dotted line showing cross section. B: schematic representations of long bone cross-sectional geometry in males vs. females at the level of the dotted line shown in A. Males typically have a wider bone cross section and outer radius (R), whereas females have a smaller outer radius but thicker bone cortex, leading to a shorter inner radius (r). As women age, bone is resorbed from the endocortical surface so that older women must increase R to compensate for an increasing r. Males have a larger R and so are at lower fracture risk even as r increases. C: increased muscle mass in males is associated with increased bone diameter, even in mice. Male mice (M) lacking myostatin show a greater increase in forelimb triceps brachii mass than female (F) knockout (KO) mice, and forelimb bone (radius) diameter is largest in M mice lacking myostatin (KO) compared with wild-type (WT) mice.
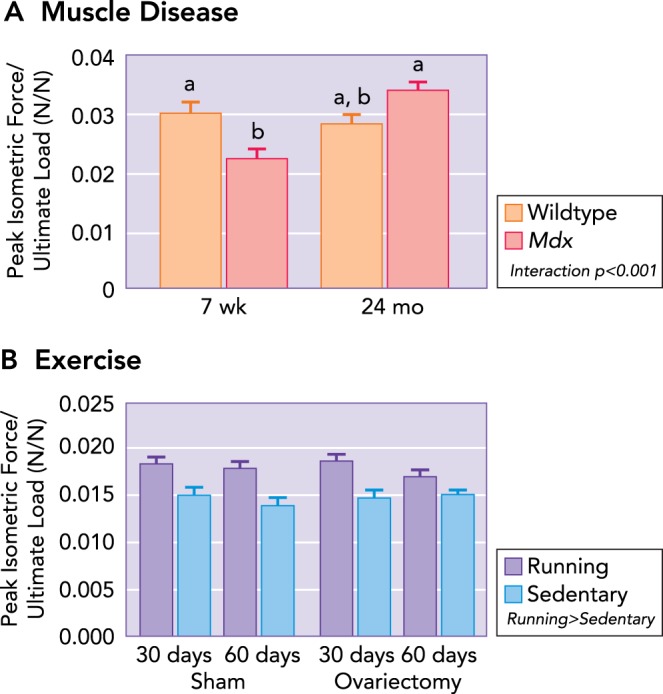
Ratio of muscle maximal isometric tetanic force to ultimate load as a function of muscle disease, physical activity, and estrogen A: at the peak of muscle-disease onset (i.e., 7 wk of age), a mismatch is present between muscle and bone, such that the bone is stronger (i.e., determined by ultimate load during three-point bending) than the adjacent musculature (i.e., strength determined by in vitro assessment of isolated muscle contractility). With aging, the muscle-to-bone ratio remains constant in healthy wild-type mice. Mdx mice become undistinguishable from 24-mo-old wild-type mice, which is attributed to an improvement in muscle function with age. Data are means (SD) and are unitless. Statistically significant effects are indicated within the figure legend. Significant interactions between genotype and age detected by Holm-Sidak post hoc tests (P < 0.05) are indicated by the lowercase letters above the bars; values with the same letter are not significantly different. B: however, the loss of estrogen does not influence the muscle-to-bone ratio; running increases the ratio between muscle and bone, suggesting a transient mismatch between muscle and bone. Values are means (SD) and are unitless. Figure was adapted with permission from Refs. , 78.
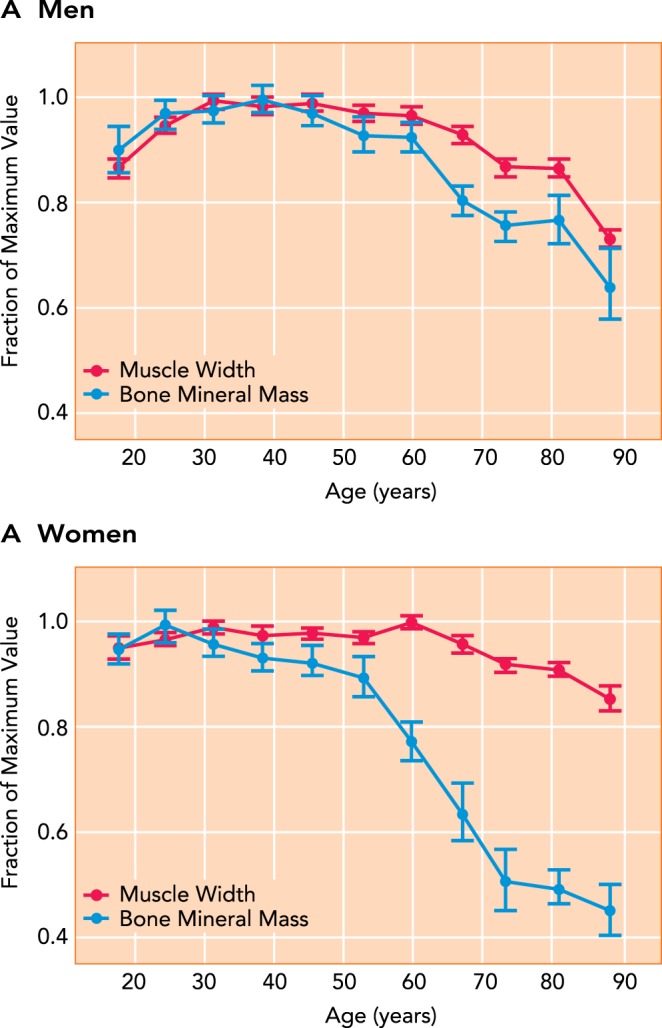
Aging-related changes in bone mineral mass of the radius and muscle width in the forearm in adult men and women The figure was adapted with permission from Ref. , and data were normalized to the peak value for bone and muscle across the lifespan to show relative changes across the lifespan for men (A) and women (B).
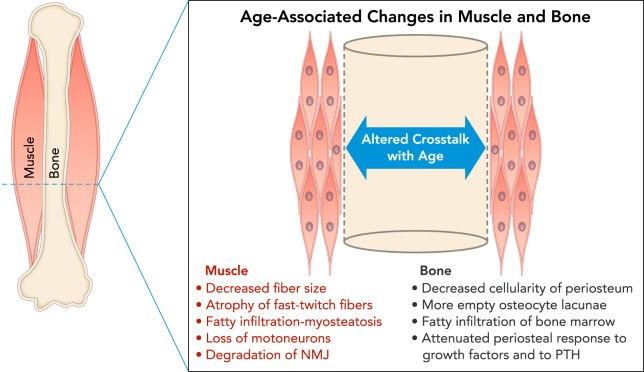
Schematic cross-section at the midshaft of a long bone diaphysis showing a summary of age-associated changes in muscle and bone Muscle changes (red) include decreased fiber size, atrophy of fast-twitch fibers, fatty infiltration of muscle tissue, loss of motoneurons, and degradation of the neuromuscular junction (NMJ), all of which can negatively impact force production and potentially the secretion of myokines. Changes in bone (black) include decreased cellularity in the periosteum, loss of osteocytes in bone matrix due to senescence, fatty infiltration of bone marrow, and attenuated periosteal response to growth factors and parathyroid hormone (PTH). All of these changes can impair the capacity of bone to respond to anabolic stimuli.
References
-
- Adams GR, Haddad F, Bodell PW, Tran PD, Baldwin KM. Combined isometric, concentric, and eccentric resistance exercise prevents unloading-induced muscle atrophy in rats. J Appl Physiol 103: 1644–1654, 2007. - PubMed
-
- Alsharidah M, Lazarus NR, George TE, Agley CC, Velloso CP, Harridge SD. Primary human muscle precursor cells obtained from young and old donors produce similar proliferative, differentiation and senescent profiles in culture. Aging Cell 12: 333–344, 2013. - PubMed
-
- Atlantis E, Martin SA, Haren MT, Taylor AW, Wittert GA. Florey Adelaide Male Aging Study. Lifestyle factors associated with age-related differences in body composition: the Florey Adelaide Male Aging Study. Am J Clin Nutr 88: 95–104, 2008. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical