

Animal model of Sar1b deficiency presents lipid absorption deficits similar to Anderson disease
- PMID: 25559265
- PMCID: PMC4319984
- DOI: 10.1007/s00109-014-1247-x
Animal model of Sar1b deficiency presents lipid absorption deficits similar to Anderson disease
Abstract
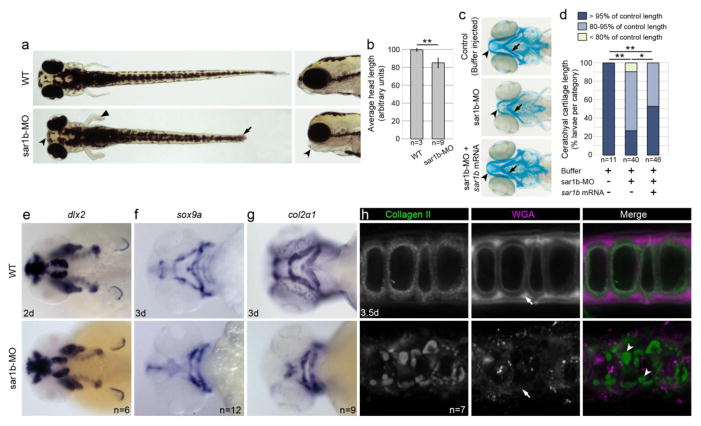
Anderson disease (ANDD) or chylomicron retention disease (CMRD) is a rare, hereditary lipid malabsorption syndrome associated with mutations in the SAR1B gene that is characterized by failure to thrive and hypocholesterolemia. Although the SAR1B structure has been resolved and its role in formation of coat protein II (COPII)-coated carriers is well established, little is known about the requirement for SAR1B during embryogenesis. To address this question, we have developed a zebrafish model of Sar1b deficiency based on antisense oligonucleotide knockdown. We show that zebrafish sar1b is highly conserved among vertebrates; broadly expressed during development; and enriched in the digestive tract organs, brain, and craniofacial skeleton. Consistent with ANDD symptoms of chylomicron retention, we found that dietary lipids in Sar1b-deficient embryos accumulate in enterocytes. Transgenic expression analysis revealed that Sar1b is required for growth of exocrine pancreas and liver. Furthermore, we found abnormal differentiation and maturation of craniofacial cartilage associated with defects in procollagen II secretion and absence of select, neuroD-positive neurons of the midbrain and hindbrain. The model presented here will help to systematically dissect developmental roles of Sar1b and to discover molecular and cellular mechanisms leading to organ-specific ANDD pathology. Key messages: Sar1b depletion phenotype in zebrafish resembles Anderson disease deficits. Sar1b deficiency results in multi-organ developmental deficits. Sar1b is required for dietary cholesterol uptake into enterocytes.
Conflict of interest statement
The authors declare that they do not have any competing or financial interests.
Figures
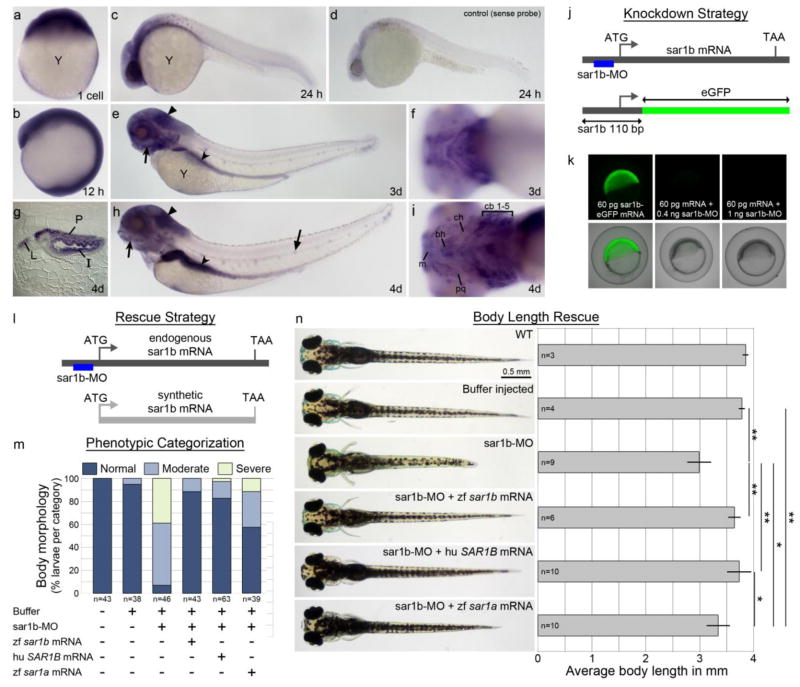
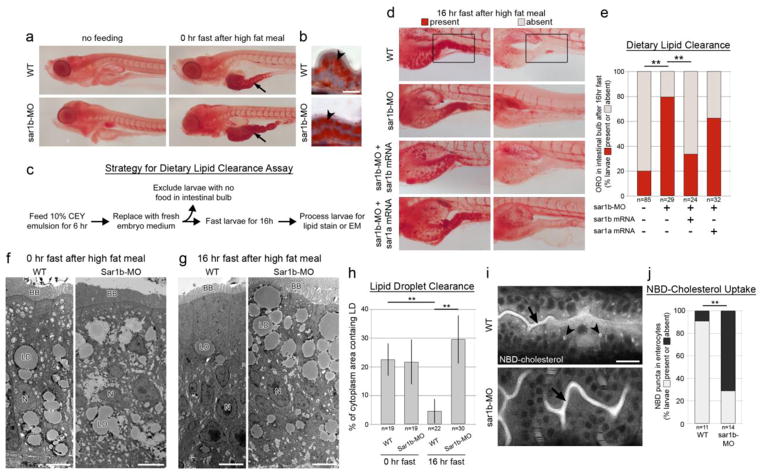
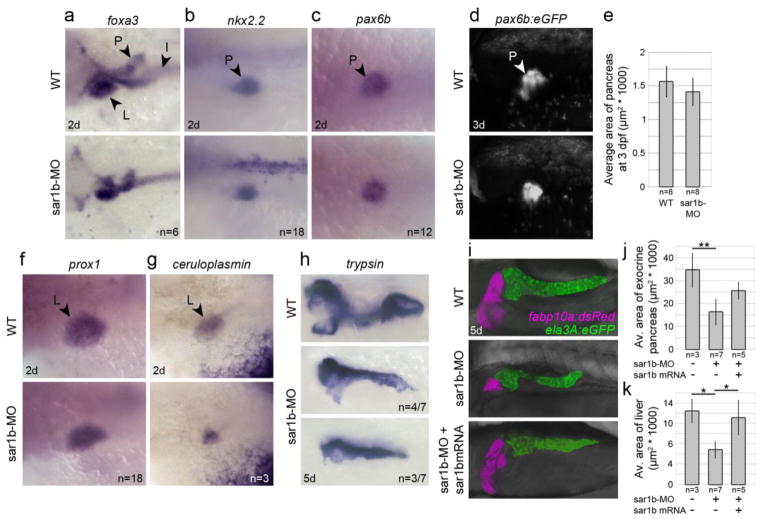
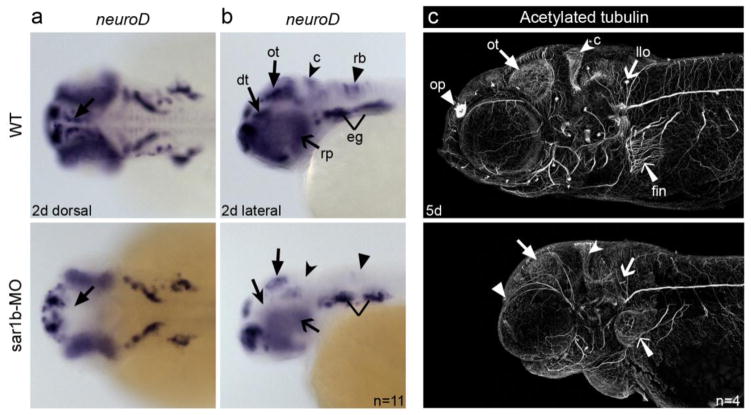
Comment in
-
Studying lipoprotein trafficking in zebrafish, the case of chylomicron retention disease.J Mol Med (Berl). 2015 Feb;93(2):115-8. doi: 10.1007/s00109-014-1248-9. J Mol Med (Berl). 2015. PMID: 25572701 No abstract available.
Similar articles
-
Anderson's disease/chylomicron retention disease in a Japanese patient with uniparental disomy 7 and a normal SAR1B gene protein coding sequence.Orphanet J Rare Dis. 2011 Nov 21;6:78. doi: 10.1186/1750-1172-6-78. Orphanet J Rare Dis. 2011. PMID: 22104167 Free PMC article.
-
Sar1b mutant mice recapitulate gastrointestinal abnormalities associated with chylomicron retention disease.J Lipid Res. 2021;62:100085. doi: 10.1016/j.jlr.2021.100085. Epub 2021 May 5. J Lipid Res. 2021. PMID: 33964306 Free PMC article.
-
Understanding Chylomicron Retention Disease Through Sar1b Gtpase Gene Disruption: Insight From Cell Culture.Arterioscler Thromb Vasc Biol. 2017 Dec;37(12):2243-2251. doi: 10.1161/ATVBAHA.117.310121. Epub 2017 Oct 5. Arterioscler Thromb Vasc Biol. 2017. PMID: 28982670
-
[Chylomicron retention disease caused by SAR1B gene variations in 2 cases and literatures review].Zhonghua Er Ke Za Zhi. 2024 Jun 2;62(6):565-570. doi: 10.3760/cma.j.cn112140-20240301-00138. Zhonghua Er Ke Za Zhi. 2024. PMID: 38763880 Review. Chinese.
-
Chylomicron retention disease: genetics, biochemistry, and clinical spectrum.Curr Opin Lipidol. 2019 Apr;30(2):134-139. doi: 10.1097/MOL.0000000000000578. Curr Opin Lipidol. 2019. PMID: 30640893 Review.
Cited by
-
Metabolic coessentiality mapping identifies C12orf49 as a regulator of SREBP processing and cholesterol metabolism.Nat Metab. 2020 Jun;2(6):487-498. doi: 10.1038/s42255-020-0206-9. Epub 2020 Jun 1. Nat Metab. 2020. PMID: 32694732 Free PMC article.
-
Studying lipoprotein trafficking in zebrafish, the case of chylomicron retention disease.J Mol Med (Berl). 2015 Feb;93(2):115-8. doi: 10.1007/s00109-014-1248-9. J Mol Med (Berl). 2015. PMID: 25572701 No abstract available.
-
Small sequence variations between two mammalian paralogs of the small GTPase SAR1 underlie functional differences in coat protein complex II assembly.J Biol Chem. 2020 Jun 19;295(25):8401-8412. doi: 10.1074/jbc.RA120.012964. Epub 2020 May 1. J Biol Chem. 2020. PMID: 32358066 Free PMC article.
-
Zebrafish Models for Dyslipidemia and Atherosclerosis Research.Front Endocrinol (Lausanne). 2016 Dec 16;7:159. doi: 10.3389/fendo.2016.00159. eCollection 2016. Front Endocrinol (Lausanne). 2016. PMID: 28018294 Free PMC article. Review.
-
Dynamic regulation of inter-organelle communication by ubiquitylation controls skeletal muscle development and disease onset.Elife. 2023 Jul 11;12:e81966. doi: 10.7554/eLife.81966. Elife. 2023. PMID: 37432316 Free PMC article.
References
-
- Siddiqi SA, Gorelick FS, Mahan JT, Mansbach CM. COPII proteins are required for Golgi fusion but not for endoplasmic reticulum budding of the pre-chylomicron transport vesicle. J Cell Sci. 2003;116:415–427. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
