

Sodium butyrate epigenetically modulates high-fat diet-induced skeletal muscle mitochondrial adaptation, obesity and insulin resistance through nucleosome positioning
- PMID: 25559882
- PMCID: PMC4439875
- DOI: 10.1111/bph.13058
Sodium butyrate epigenetically modulates high-fat diet-induced skeletal muscle mitochondrial adaptation, obesity and insulin resistance through nucleosome positioning
Abstract
Background and purpose: Sodium butyrate (NaB), an epigenetic modifier, is effective in promoting insulin sensitivity. The specific genomic loci and mechanisms underlying epigenetically induced obesity and insulin resistance and the targets of NaB are not fully understood.
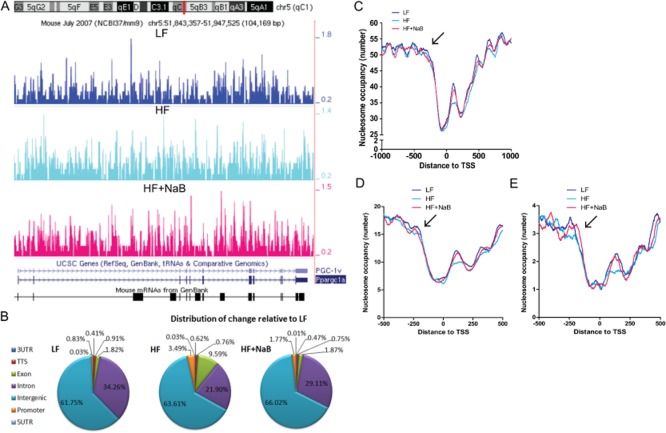
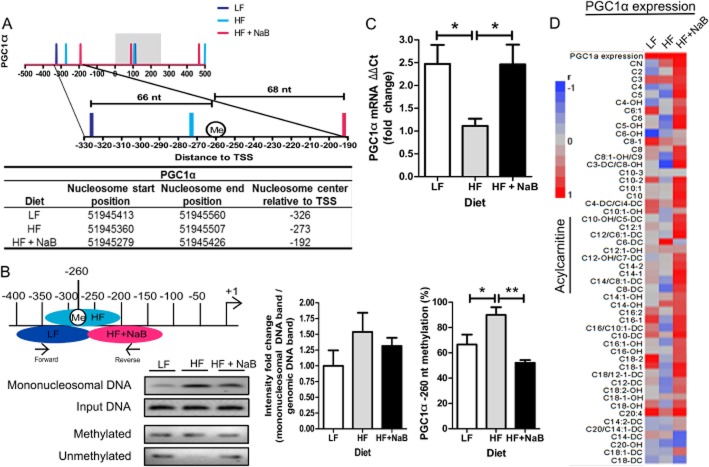
Experimental approach: The anti-diabetic and anti-obesity effects of NaB treatment were measured by comparing phenotypes and physiologies of C57BL/6J mice fed a low-fat diet (LF), high-fat diet (HF) or high-fat diet plus NaB (HF + NaB) for 10 weeks. We determined a possible mechanism of NaB action through induction of beneficial skeletal muscle mitochondrial adaptations and applied microccocal nuclease digestion with sequencing (MNase-seq) to assess whole genome differences in nucleosome occupancy or positioning and to identify associated epigenetic targets of NaB.
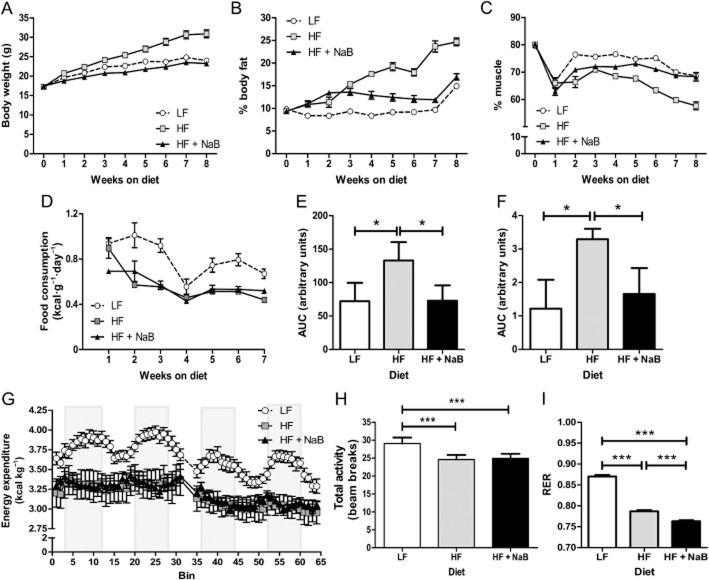
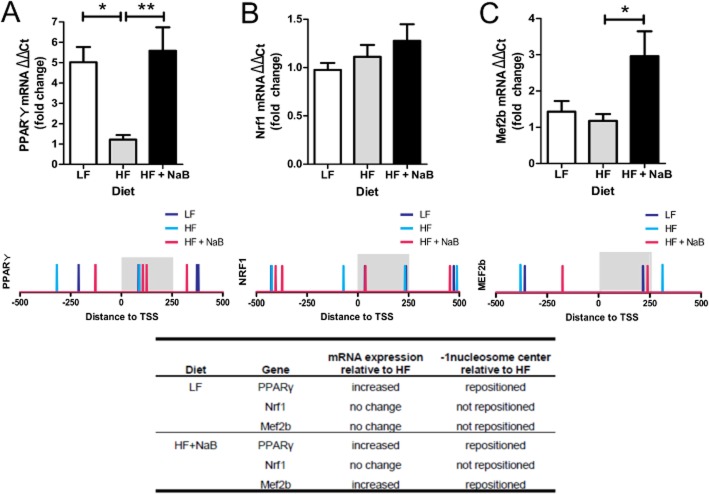
Key results: NaB prevented HF diet-induced increases in body weight and adiposity without altering food intake or energy expenditure, improved insulin sensitivity as measured by glucose and insulin tolerance tests, and decreased respiratory exchange ratio. In skeletal muscle, NaB increased the percentage of type 1 fibres, improved acylcarnitine profiles as measured by metabolomics and produced a chromatin structure, determined by MNase-seq, similar to that seen in LF. Targeted analysis of representative nuclear-encoded mitochondrial genes showed specific repositioning of the -1 nucleosome in association with altered gene expression.
Conclusions and implications: NaB treatment may be an effective pharmacological approach for type 2 diabetes and obesity by inducing -1 nucleosome repositioning within nuclear-encoded mitochondrial genes, causing skeletal muscle mitochondrial adaptations that result in more complete β-oxidation and a lean, insulin sensitive phenotype.
© 2015 The British Pharmacological Society.
Figures
References
-
- Albert I, Mavrich TN, Tomsho LP, Qi J, Zanton SJ, Schuster SC, et al. Translational and rotational settings of H2A.Z nucleosomes across the Saccharomyces cerevisiae genome. Nature. 2007;446:572–576. - PubMed
-
- Barrès R, Osler ME, Yan J, Rune A, Fritz T, Caidahl K, et al. Non-CpG methylation of the PGC-1α promoter through DNMT3B controls mitochondrial density. Cell Metab. 2009;10:189–198. - PubMed
-
- Barrès R, Yan J, Egan B, Treebak Jonas T, Rasmussen M, Fritz T, et al. Acute exercise remodels promoter methylation in human skeletal muscle. Cell Metab. 2012;15:405–411. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
