

Mechanisms of retinoic acid signalling and its roles in organ and limb development
- PMID: 25560970
- PMCID: PMC4636111
- DOI: 10.1038/nrm3932
Mechanisms of retinoic acid signalling and its roles in organ and limb development
Abstract
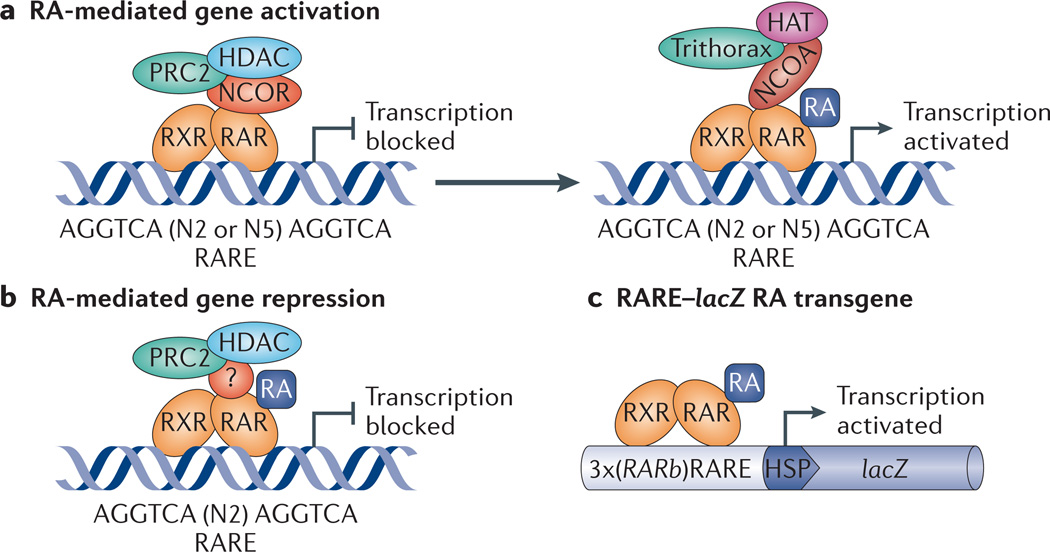
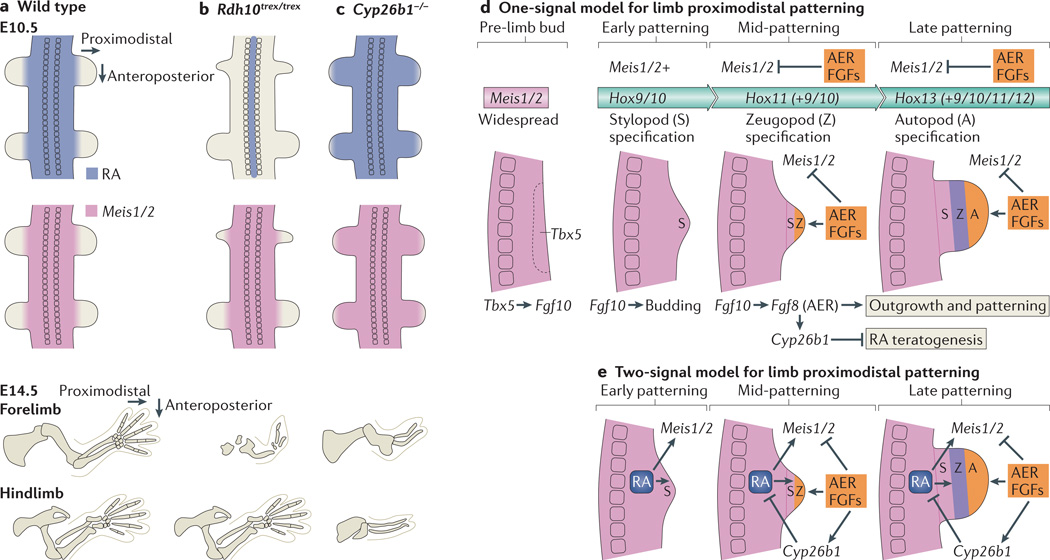
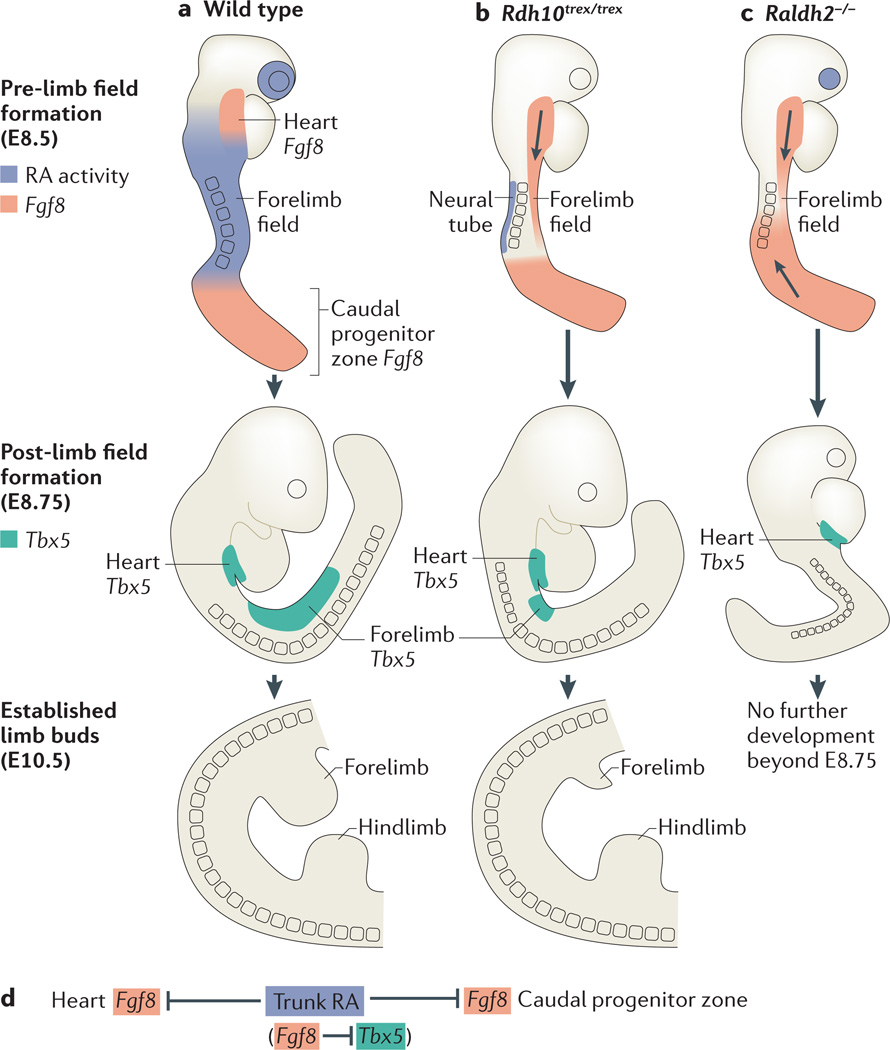
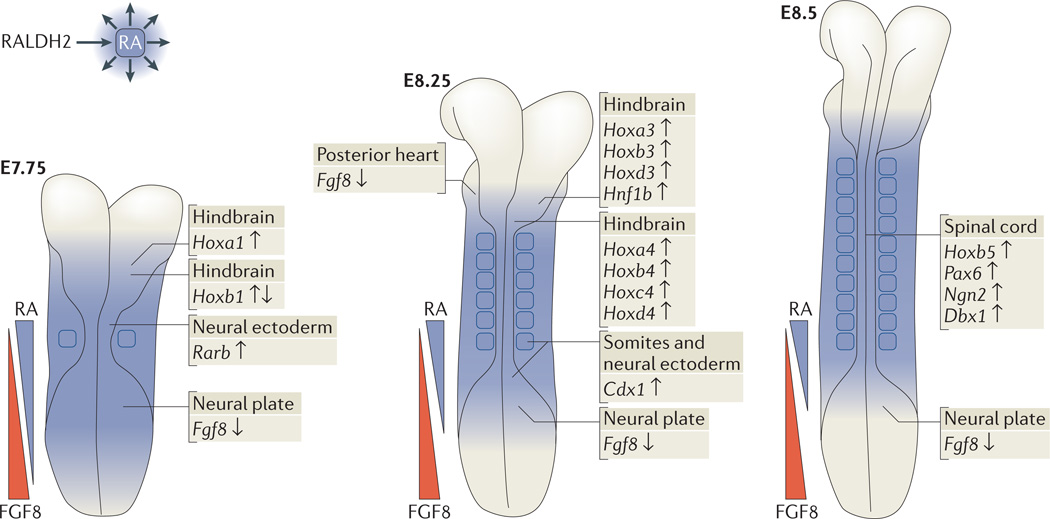
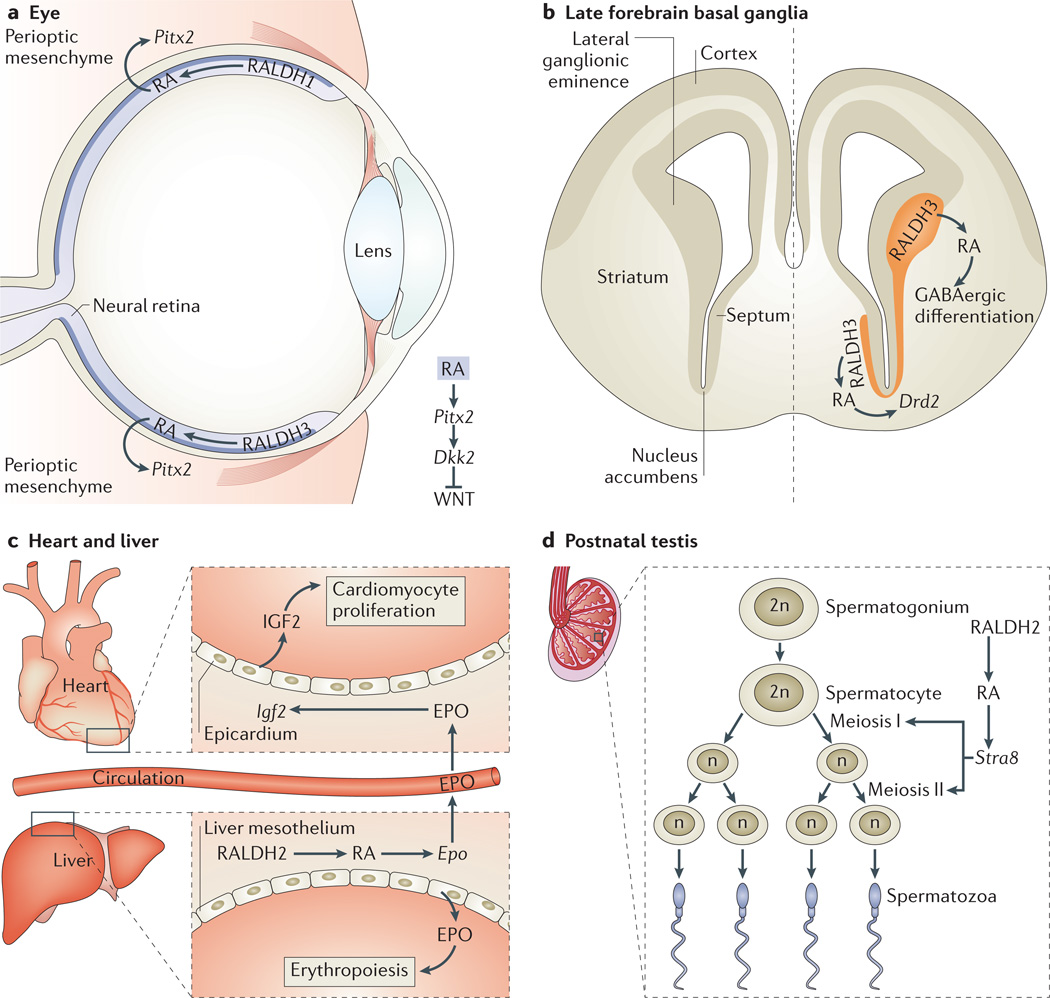
Retinoic acid (RA) signalling has a central role during vertebrate development. RA synthesized in specific locations regulates transcription by interacting with nuclear RA receptors (RARs) bound to RA response elements (RAREs) near target genes. RA was first implicated in signalling on the basis of its teratogenic effects on limb development. Genetic studies later revealed that endogenous RA promotes forelimb initiation by repressing fibroblast growth factor 8 (Fgf8). Insights into RA function in the limb serve as a paradigm for understanding how RA regulates other developmental processes. In vivo studies have identified RAREs that control repression of Fgf8 during body axis extension or activation of homeobox (Hox) genes and other key regulators during neuronal differentiation and organogenesis.
Figures
References
-
- Frazier CN, Hu CK. Cutaneous lesions associated with a deficiency in vitamin A in man. Arch. Intern. Med. 1931;48:507–514.
-
- Wilson JG, Roth CB, Warkany J. An analysis of the syndrome of malformations induced by maternal vitamin A deficiency. Effects of restoration of vitamin A at various times during gestation. Amer. J. Anat. 1953;92:189–217. - PubMed
-
- Clagett-Dame M, DeLuca HF. The role of vitamin A in mammalian reproduction and embryonic development. Annu. Rev. Nutr. 2002;22:347–381. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
