

Scl binds to primed enhancers in mesoderm to regulate hematopoietic and cardiac fate divergence
- PMID: 25564442
- PMCID: PMC4369313
- DOI: 10.15252/embj.201490542
Scl binds to primed enhancers in mesoderm to regulate hematopoietic and cardiac fate divergence
Abstract
Scl/Tal1 confers hemogenic competence and prevents ectopic cardiomyogenesis in embryonic endothelium by unknown mechanisms. We discovered that Scl binds to hematopoietic and cardiac enhancers that become epigenetically primed in multipotent cardiovascular mesoderm, to regulate the divergence of hematopoietic and cardiac lineages. Scl does not act as a pioneer factor but rather exploits a pre-established epigenetic landscape. As the blood lineage emerges, Scl binding and active epigenetic modifications are sustained in hematopoietic enhancers, whereas cardiac enhancers are decommissioned by removal of active epigenetic marks. Our data suggest that, rather than recruiting corepressors to enhancers, Scl prevents ectopic cardiogenesis by occupying enhancers that cardiac factors, such as Gata4 and Hand1, use for gene activation. Although hematopoietic Gata factors bind with Scl to both activated and repressed genes, they are dispensable for cardiac repression, but necessary for activating genes that enable hematopoietic stem/progenitor cell development. These results suggest that a unique subset of enhancers in lineage-specific genes that are accessible for regulators of opposing fates during the time of the fate decision provide a platform where the divergence of mutually exclusive fates is orchestrated.
Keywords: cardiac specification; enhancer; hematopoiesis; mesoderm diversification; transcriptional regulation.
© 2015 The Authors.
Figures
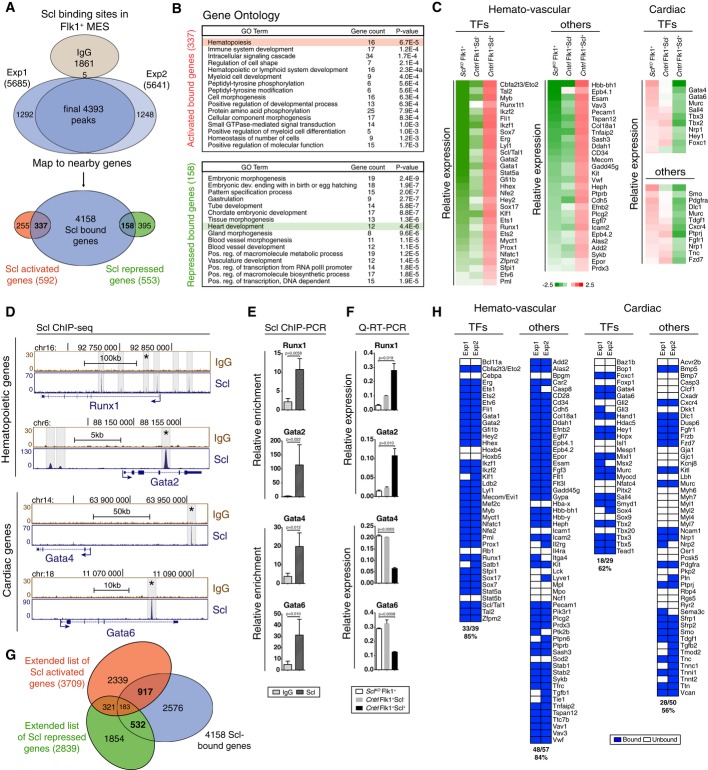
Venn diagram showing the number of Scl binding sites and overlap with Scl activated and repressed genes in Flk1+ MES (mesoderm) documents that Scl binds to both Scl-dependent activated and repressed genes.
DAVID (Huang et al, 2007) GO enrichment analysis for Scl-bound and activated or Scl-bound and repressed genes shows enrichment of hematopoietic and heart-related terms, respectively.
Gene expression heatmaps of selected genes from the bound and activated and bound and repressed groups show activation of key hematopoietic target genes and repression of key cardiac target genes in Scl-expressing mesoderm as compared to Scl-deficient mesoderm. Control [Scl+/hCD4, divided into Scl-expressing (Scl+) and non-expressing cells (Scl−)] and SclKO EBs are shown.
Scl and control IgG ChIP-seq tracks show examples of Scl binding sites near hematopoietic (Runx1 and Gata2) and cardiac (Gata4 and Gata6) genes in Flk1+ mesoderm (MES).
Verification of Scl binding sites marked with asterisks in (D) using ChIP-PCR, average enrichment of at least three independent biological experiments over negative control region (chr16: 92230219–92230338) with SEM are shown.
Verification of Scl-dependent gene expression using qRT–PCR. Average of three biological replicates with SD are shown.
Venn diagrams show the number of potential ‘extended activated’ and ‘extended repressed’ Scl target genes by intersecting genes associated with Scl binding in Flk1+ MES with Scl activated and repressed genes from day 4 EBs, E9.5 yolk sac, placenta and endocardium (Van Handel et al, 2012).
Heatmaps show Scl binding to majority of activated hematopoietic and repressed cardiac transcription factors and other proteins. See also Supplementary Fig S1 and Supplementary Table S1A–E.
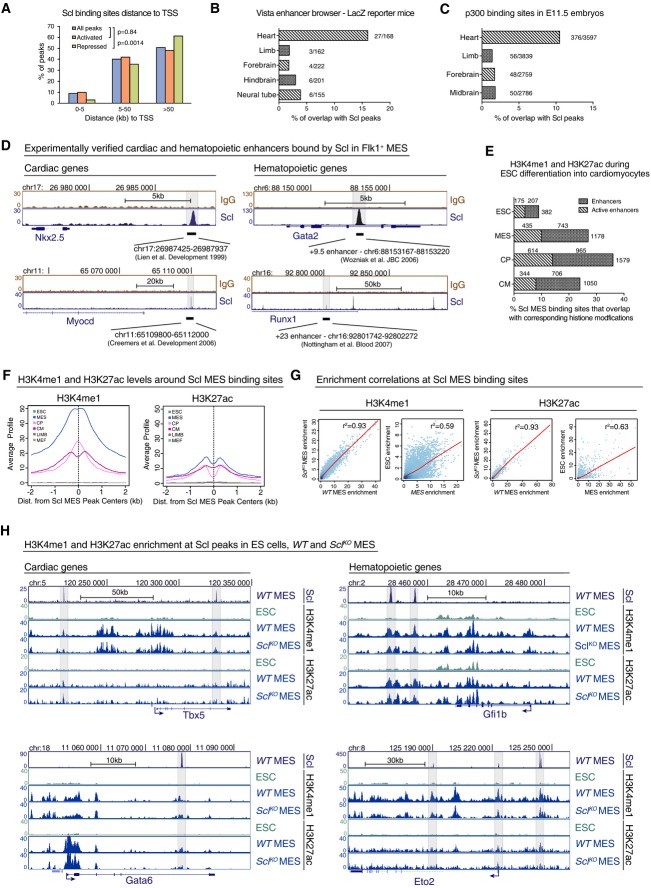
Distribution of Scl binding sites relative to TSS in Scl-bound activated or repressed genes shows that majority of Scl binding sites locate away from TSS. Chi-square test was used to assess differences in the distribution.
Percent of tissue-specific enhancers validated by LacZ reporter mice (Vista enhancer browser) that overlap with Scl MES binding sites shows that Scl binds to experimentally verified heart enhancers more often compared to enhancers from other tissues.
Percent of Scl MES binding sites that overlap with heart enhancers in E11.5 embryos defined by p300 binding (Blow et al, 2010) is higher than overlap with enhancers from other tissues.
Scl MES binding sites overlap with experimentally verified heart-specific enhancers upstream of Myocardin and Nkx2.5 genes and with hematopoietic enhancers within Runx1 and Gata2 genes.
Percent of enhancers from different cardiac developmental stages identified by H3K4me1 (enhancers) and H3K4me1 combined with H3k27ac (active enhancers) (Wamstad et al, 2012) that overlap with Scl MES binding sites.
Average H3K4me1 (left) and H3K27ac (right) profiles around all 4,393 Scl binding sites show tissue-specific enrichment in Flk1+ mesoderm (MES), cardiac precursors (CP) and cardiomyocytes (CM) but not in ESC (mouse ES cells) mouse embryonic fibroblasts (MEF) and embryonic limbs (LIMB).
Correlation analysis of H3K4me1 (left) and H3K27ac (right) levels between WT and SclKO MES and ES cells and MES (Wamstad et al, 2012) around 4,393 Scl binding sites shows that establishment of these marks occurs independently of Scl.
Scl, H3K4me1 and H3K27ac ChIP-seq tracks show comparable levels of H3K4me1 and H3K27ac in WT and SclKO mesoderm around cardiac (Gata6, Tbx5) and hematopoietic (Eto2, Gfi1b) genes.
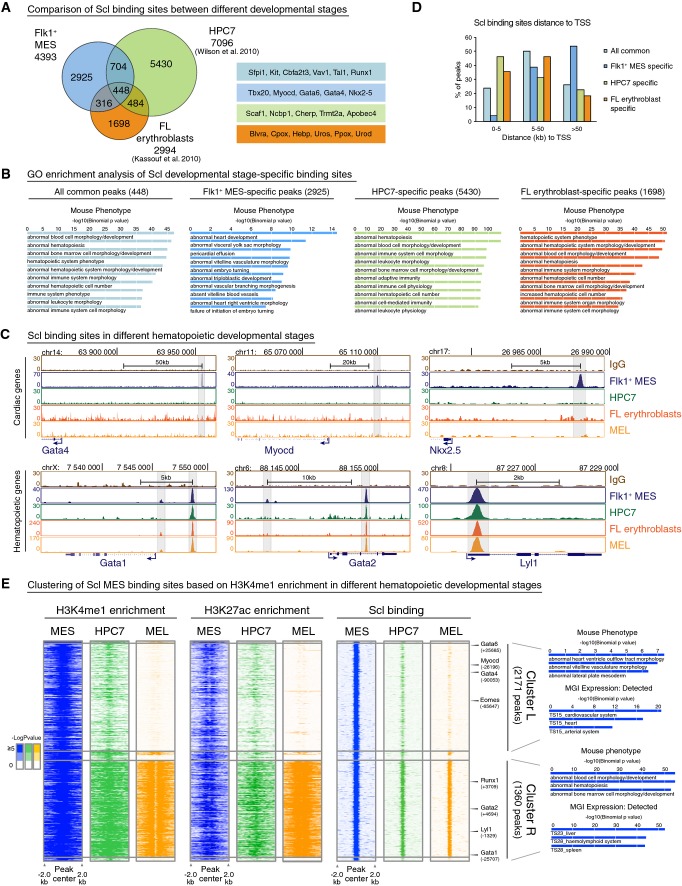
Venn diagram showing overlaps of Scl binding sites in different developmental stages—Flk1+ MES, fetal liver (FL) erythroblast cells (Kassouf et al, 2010) and hematopoietic progenitors (HPC7) (Wilson et al, 2010) (left)—shows that majority of Scl binding sites are developmental stage specific. Examples of developmental stage-specific Scl-bound genes are shown (right).
GREAT (McLean et al, 2010) analysis of significantly enriched GO terms in Mouse Phenotype category for all common, Flk1+ MES-specific, HPC7-specific and FL erythroblast-specific peaks shows enrichment of heart-related terms only among Flk1+ MES-specific peaks.
ChIP-seq tracks with Scl binding in Flk1+ MES, HPC7, FL erythroblasts and MEL (mouse erythroleukemia) cells around cardiac (Gata4, Myocd, Nkx2.5) and hematopoietic (Gata1, Gata2 and Lyl1) genes show that binding to cardiac genes occurs only in mesoderm, while binding to hematopoietic genes is maintained throughout hematopoietic development.
Comparison of the location of developmental stage-specific Scl binding sites relative to TSS shows that Scl binding near TSS occurs more often in later hematopoietic development compared to mesoderm.
Heatmaps of combinatorial clustering of H3K4me1 around Scl mesodermal binding sites in MES, HPC7 and MEL reveal two major clusters: Cluster L loses H3K4me1, H3K27ac and Scl binding during hematopoietic differentiation and is enriched for genes involved in heart development; cluster R retains H3K4me1, H3K27ac and Scl binding during hematopoietic differentiation and is enriched for hematopoiesis-related genes. Selected GO terms among top 10 most enriched are shown.
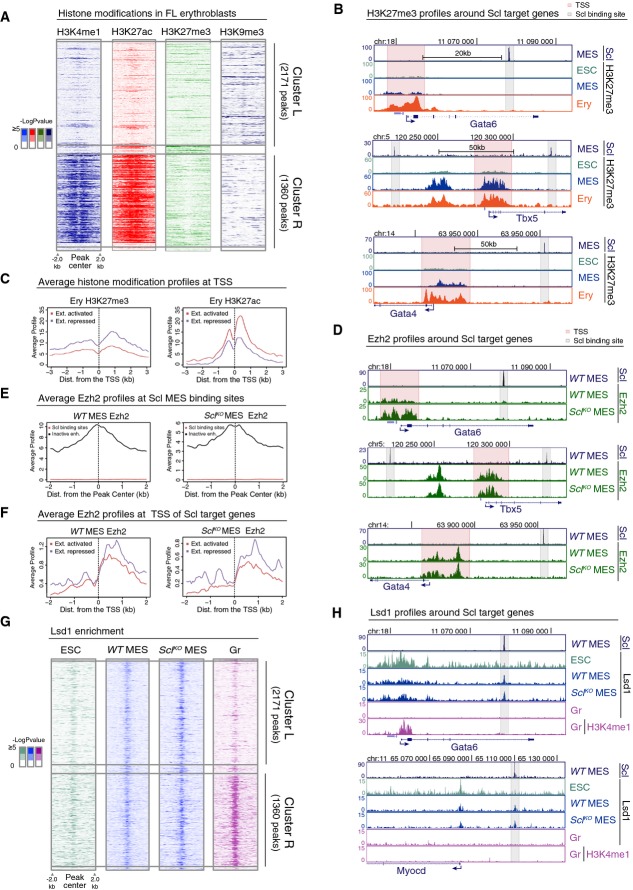
Heatmaps of H3K4me1, H3K27ac, H3K27me3 and H3K9me3 histone modifications in FL erythroblasts around Scl MES binding sites in cluster L and cluster R show no gain of common repressive histone marks in cluster L.
H3K27me3 ChIP-seq tracks in ES cells (ESC), Flk1+ mesoderm (MES) and FL erythroblasts (Ery) show that cardiac genes Gata6, Tbx5 and Gata4 harbor H3K27me3 at the TSS (pink) but not at Scl binding sites (gray).
Average H3K27me3 (left) and H3K27ac (right) levels in FL erythroblasts around the TSS of extended list of Scl activated and repressed genes show that Scl repressed genes have on average more H3K27me3 and less H3K27ac as compared to Scl activated genes.
Ezh2 ChIP-seq tracks in WT and SclKO mesoderm showing that Ezh2 recruitment to the TSS of cardiac genes Gata6, Tbx5 and Gata4 is not Scl dependent.
Average Ezh2 enrichment in WT (left) and SclKO mesoderm (right) for inactive enhancers (defined by having H3K4me1 and H3K27me3 modifications (Wamstad et al, 2012) and distance greater than 5 kb from TSS) and for Scl binding sites shows no Ezh2 enrichment at Scl binding sites.
Average Ezh2 enrichment at the TSS of extended list of Scl activated and repressed genes is similar in WT (left) and SclKO mesoderm (right).
Heatmaps of Lsd1 enrichment in ES cells, WT and SclKO mesoderm and in granulocytes (Gr) (Kerenyi et al, 2013) in cluster L and cluster R show co-localization of Lsd1 binding with Scl binding sites and reveal that recruitment of Lsd1 can happen independently of Scl.
Lsd1 ChIP-seq tracks (ES cells, WT and SclKO Flk1+ mesoderm and granulocytes) and H3K4me1 ChIP-seq track (granulocytes) show that Lsd1 enrichment coincides with Scl MES binding sites nearby cardiac genes Gata6 and Myocd in WT and SclKO Flk1+ mesoderm.
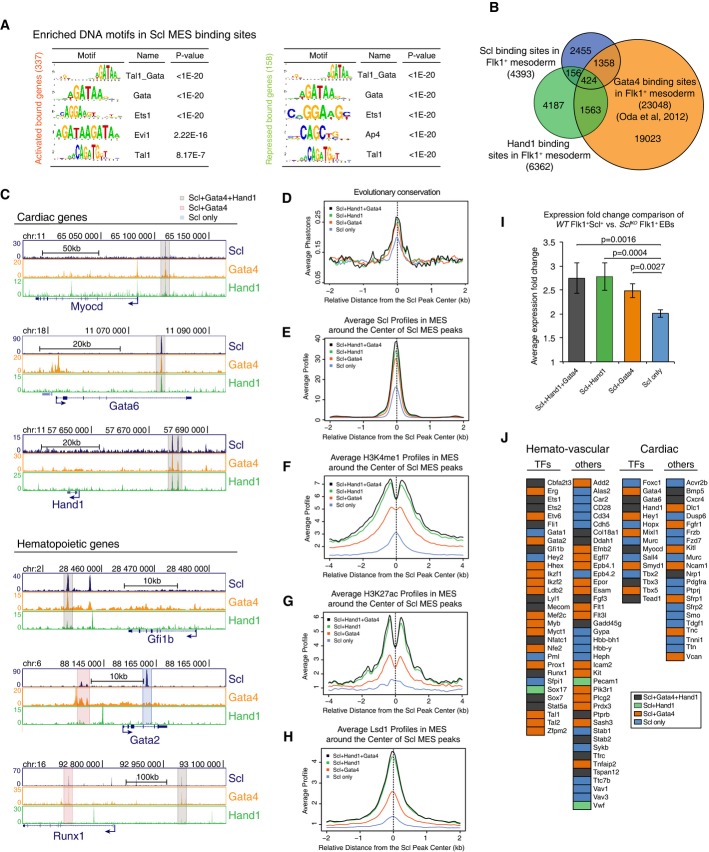
A Scl DNA binding motif analysis shows enrichment of Tal1_Gata, Gata and Ets motifs in both Scl activated and repressed genes.
B Venn diagram showing overlaps between Scl, Hand1 and Gata4 (Oda et al, 2013) binding sites in Flk1+ mesoderm documents that some of Scl MES binding sites are also bound by cardiac TFs.
C Gata4, Hand1 and Scl ChIP-seq tracks with cardiac and hematopoietic gene regions show examples of three subgroups of enhancers—those bound by Scl, Hand1 and Gata4 (gray), Scl and Gata4 (orange) and those bound only by Scl (blue).
D-H Scl binding sites that overlap with both Hand1 and Gata4 (gray) or with Hand1 (green) or Gata 4 (orange) alone show higher evolutionary conservation (D) and higher average enrichment of Scl (E), H3K4me1 (F), H3K27ac (G) and Lsd1 (H) in mesoderm compared to sites bound by Scl alone (blue).
I Average expression changes between Scl-expressing and Scl-deficient mesoderm shows that genes that are regulated by enhancers bound by all three (Scl, Hand1 and Gata4) factors or two factors (Scl and Hand1 or Scl and Gata4) show higher absolute expression changes compared to those bound by Scl alone.
J Analysis of Scl-regulated key hematopoietic and cardiac genes shows that many of them can be bound also by cardiac factors, Scl, Gata4 and Hand1 (orange); Scl and Hand1 binding (green); Scl and Gata4 binding (orange); and Scl binding alone (blue).
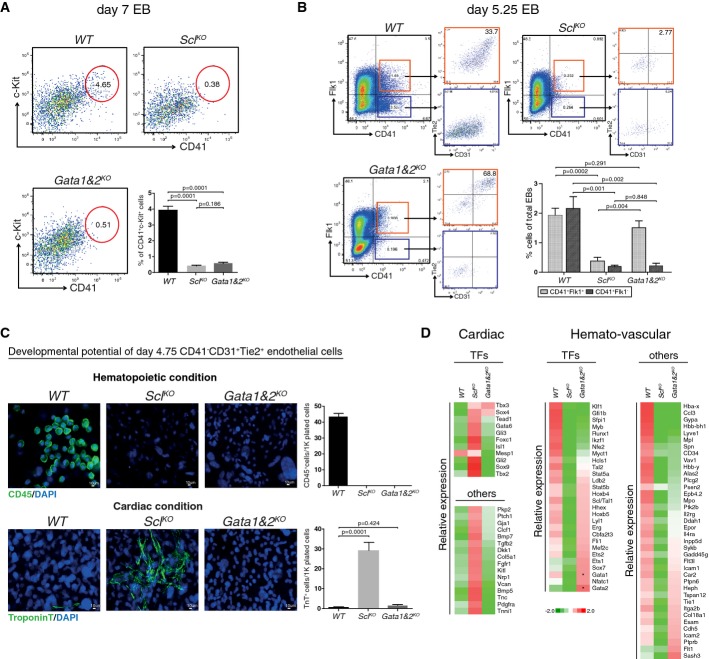
FACS analysis of day 7 EBs with markers CD41 and c-Kit shows efficient generation of HS/PCs from WT, but not SclKO or Gata1&2KO cells. Average of six independent experiments with SEM is shown.
FACS analysis of day 5.25 EBs with markers CD41, Flk1, Tie2 and CD31 shows the generation of hemogenic endothelial cells from WT and Gata1&2KO cells, but not SclKO cells. Average of five independent biological experiments with SEM is shown.
Assessment of the developmental potential of day 4.75 EB CD41−CD31+Tie2+ endothelial cells on OP9 for 14 days shows the generation of CD45+ hematopoietic cells from WT cells but not Gata1&2KO or SclKO cells, and troponin T+ cardiomyocytes from SclKO cells, but not WT or Gata1&2KO cells. Average of four independent experiments with SEM is shown.
Heatmaps show gene expression differences in subsets of hematopoietic genes between Gata1&2KO and WT or SclKO day 4.75 CD41−CD31+Tie2+ endothelial cells. Cardiac derepression is observed only in SclKO cells. *designates non-functional transcripts.
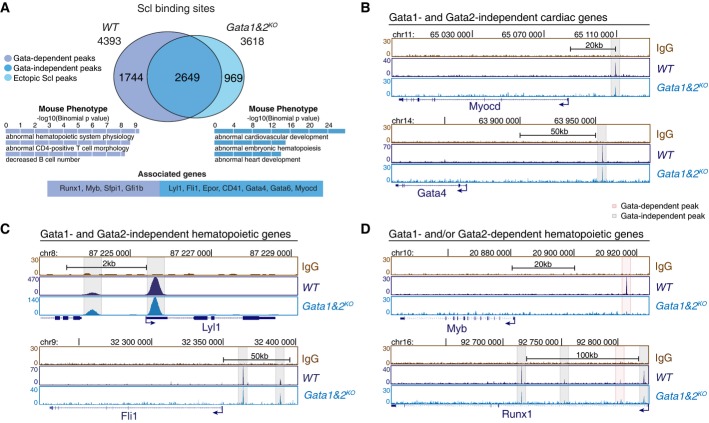
A Intersection of Scl binding sites in WT and Gata1&2KO Flk1+ MES shows Gata1- and/or Gata2-dependent redistribution of Scl binding sites in a subset of target genes (two independent experiments). Selected GO terms among top 10 most enriched for Gata-independent (blue) and Gata-dependent (purple) Scl binding sites together with example genes reveal Gata1- and/or Gata2-dependent functions for Scl-mediated hematopoietic regulation.
B-D Scl ChIP-seq tracks in WT and Gata1&2KO Flk1+ MES show Gata1- and 2-independent Scl binding to cardiac regulators (Myocardin, Gata4) (B), Gata1- and 2-independent Scl binding to key hemato-vascular regulators (Lyl1, Fli1) (C) and Gata1- and/or 2-dependent Scl binding to key HS/PC transcription factors (Myb, Runx1) (D).
Comment in
-
Creating cellular diversity through transcription factor competition.EMBO J. 2015 Mar 12;34(6):691-3. doi: 10.15252/embj.201591017. Epub 2015 Feb 13. EMBO J. 2015. PMID: 25680687 Free PMC article.
References
-
- Amente S, Lania L, Majello B. The histone LSD1 demethylase in stemness and cancer transcription programs. Biochim Biophys Acta. 2013;1829:981–986. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
