

Gain of Olig2 function in oligodendrocyte progenitors promotes remyelination
- PMID: 25564492
- PMCID: PMC4441088
- DOI: 10.1093/brain/awu375
Gain of Olig2 function in oligodendrocyte progenitors promotes remyelination
Abstract
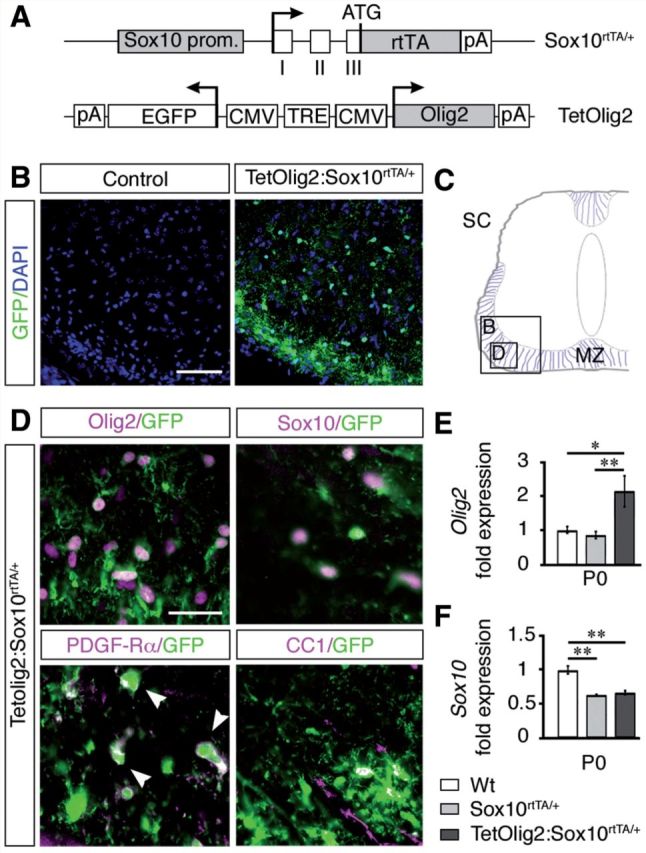
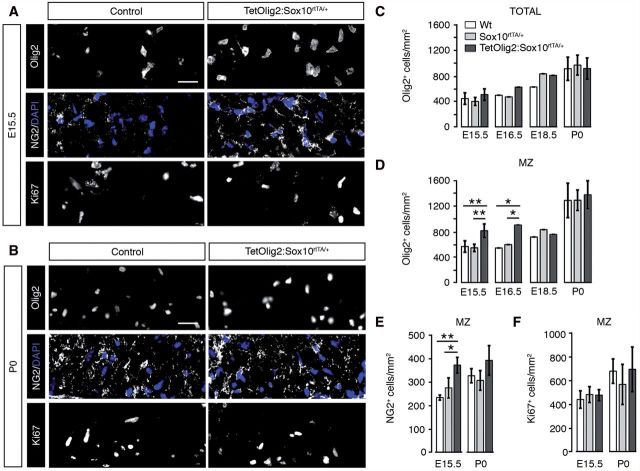
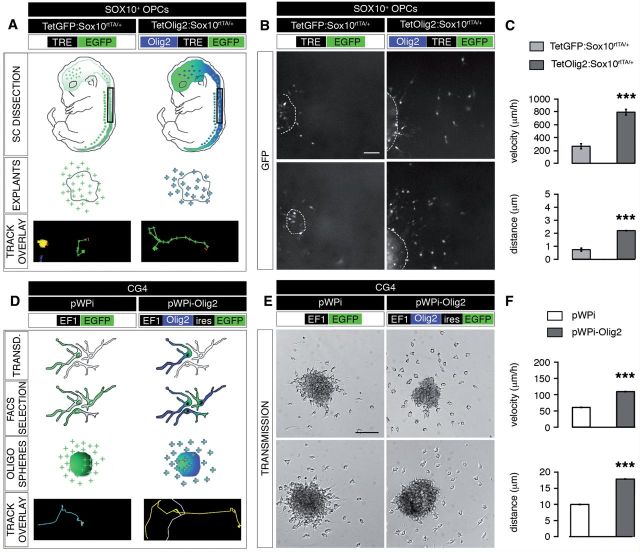
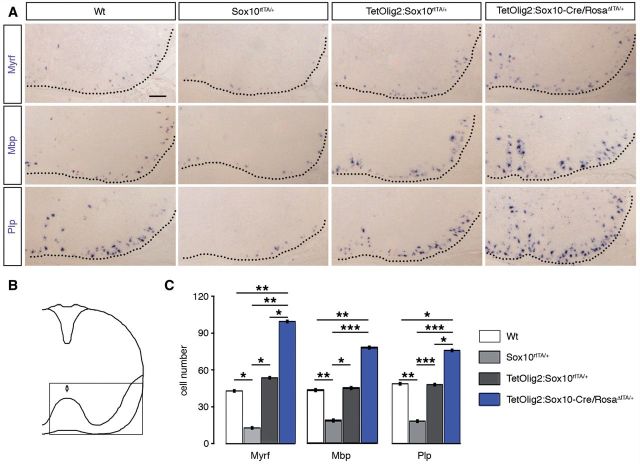
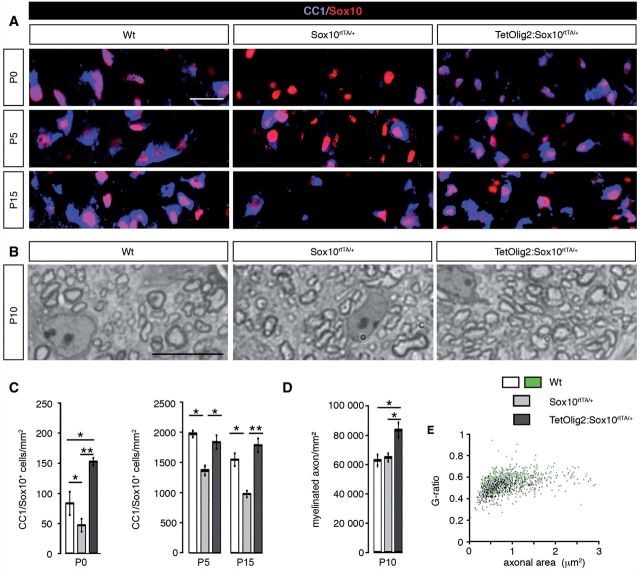
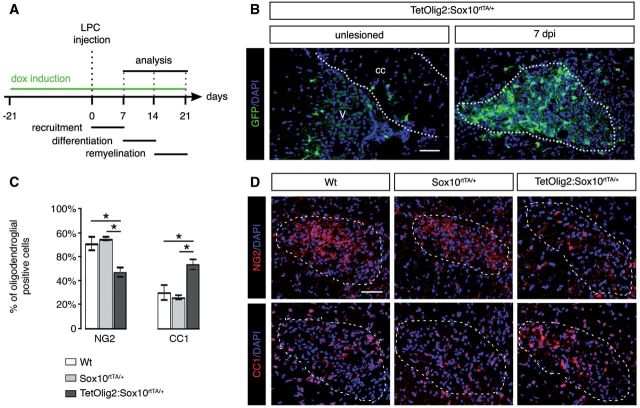
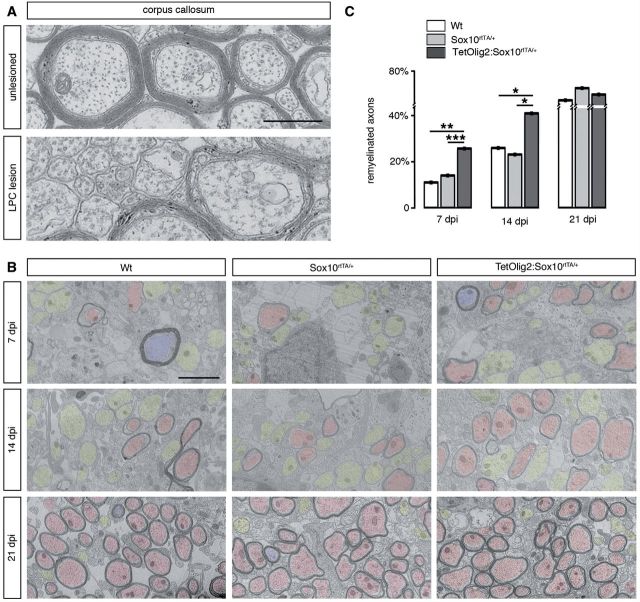
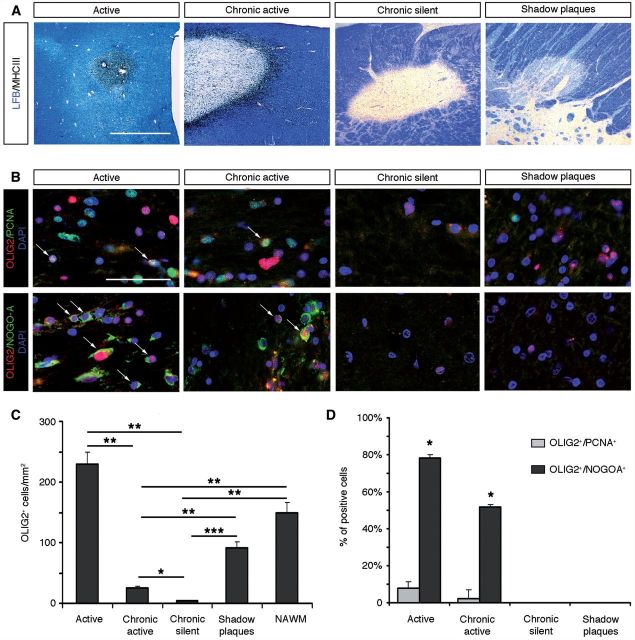
The basic helix-loop-helix transcription factor Olig2 is a key determinant for the specification of neural precursor cells into oligodendrocyte progenitor cells. However, the functional role of Olig2 in oligodendrocyte migration and differentiation remains elusive both during developmental myelination and under demyelinating conditions of the adult central nervous system. To decipher Olig2 functions, we generated transgenic mice (TetOlig2:Sox10(rtTA/+)) overexpressing Olig2 in Sox10(+) oligodendroglial cells in a doxycycline inducible manner. We show that Olig2 overexpression increases the generation of differentiated oligodendrocytes, leading to precocious myelination of the central nervous system. Unexpectedly, we found that gain of Olig2 function in oligodendrocyte progenitor cells enhances their migration rate. To determine whether Olig2 overexpression in adult oligodendrocyte progenitor cells promotes oligodendrocyte regeneration for myelin repair, we induced lysophosphatidylcholine demyelination in the corpus callosum of TetOlig2:Sox10(rtTA/+) and control mice. We found that Olig2 overexpression enhanced oligodendrocyte progenitor cell differentiation and remyelination. To assess the relevance of these findings in demyelinating diseases, we also examined OLIG2 expression in multiple sclerosis lesions. We demonstrate that OLIG2 displays a differential expression pattern in multiple sclerosis lesions that correlates with lesion activity. Strikingly, OLIG2 was predominantly detected in NOGO-A(+) (now known as RTN4-A) maturing oligodendrocytes, which prevailed in active lesion borders, rather than chronic silent and shadow plaques. Taken together, our data provide proof of principle indicating that OLIG2 overexpression in oligodendrocyte progenitor cells might be a possible therapeutic mechanism for enhancing myelin repair.
Keywords: Olig2; multiple sclerosis; oligodendrocyte; remyelination; tetracycline system.
© The Author (2014). Published by Oxford University Press on behalf of the Guarantors of Brain. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures
References
-
- Arnett HA, Fancy SP, Alberta JA, Zhao C, Plant SR, Kaing S, et al. bHLH transcription factor Olig1 is required to repair demyelinated lesions in the CNS. Science. 2004;306:2111–15. - PubMed
-
- Avellana-Adalid V, Nait-Oumesmar B, Lachapelle F, Baron-Van Evercooren A. Expansion of rat oligodendrocyte progenitors into proliferative “oligospheres” that retain differentiation potential. J Neurosci Res. 1996;45:558–70. - PubMed
-
- Chang A, Tourtellotte WW, Rudick R, Trapp BD. Premyelinating oligodendrocytes in chronic lesions of multiple sclerosis. N Engl J Med. 2002;346:165–73. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
