

Lysine demethylase KDM4A associates with translation machinery and regulates protein synthesis
- PMID: 25564516
- PMCID: PMC4355328
- DOI: 10.1158/2159-8290.CD-14-1326
Lysine demethylase KDM4A associates with translation machinery and regulates protein synthesis
Abstract
Chromatin-modifying enzymes are predominantly nuclear; however, these factors are also localized to the cytoplasm, and very little is known about their role in this compartment. In this report, we reveal a non-chromatin-linked role for the lysine-specific demethylase KDM4A. We demonstrate that KDM4A interacts with the translation initiation complex and affects the distribution of translation initiation factors within polysome fractions. Furthermore, KDM4A depletion reduced protein synthesis and enhanced the protein synthesis suppression observed with mTOR inhibitors, which paralleled an increased sensitivity to these drugs. Finally, we demonstrate that JIB-04, a JmjC demethylase inhibitor, suppresses translation initiation and enhances mTOR inhibitor sensitivity. These data highlight an unexpected cytoplasmic role for KDM4A in regulating protein synthesis and suggest novel potential therapeutic applications for this class of enzyme.
Significance: This report documents an unexpected cytoplasmic role for the lysine demethylase KDM4A. We demonstrate that KDM4A interacts with the translation initiation machinery, regulates protein synthesis and, upon coinhibition with mTOR inhibitors, enhances the translation suppression and cell sensitivity to these therapeutics.
©2015 American Association for Cancer Research.
Conflict of interest statement
Conflict of Interest: The authors declare competing financial interests because JRW is a consultant for QSonica.
Figures
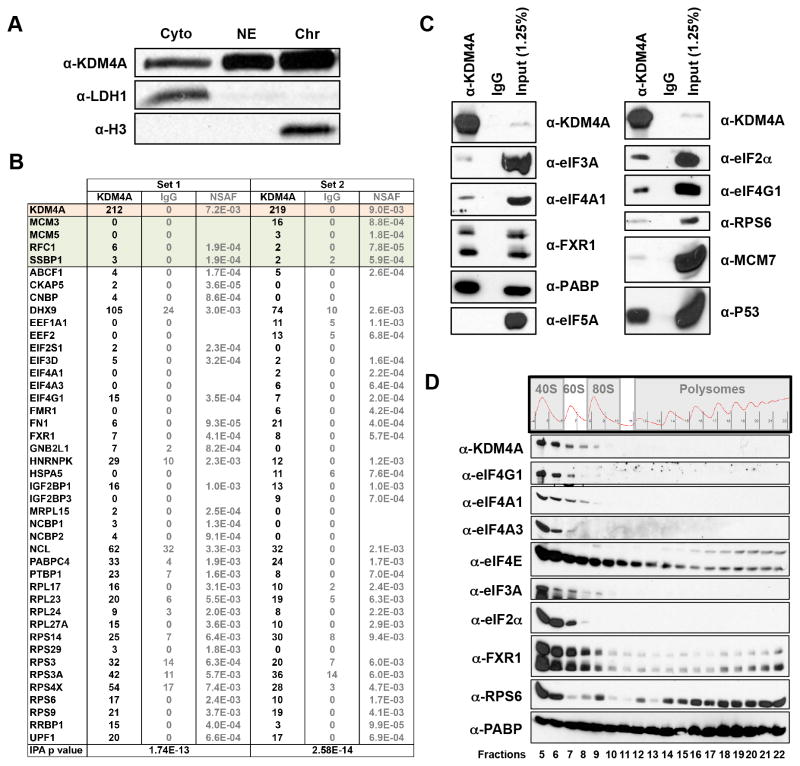
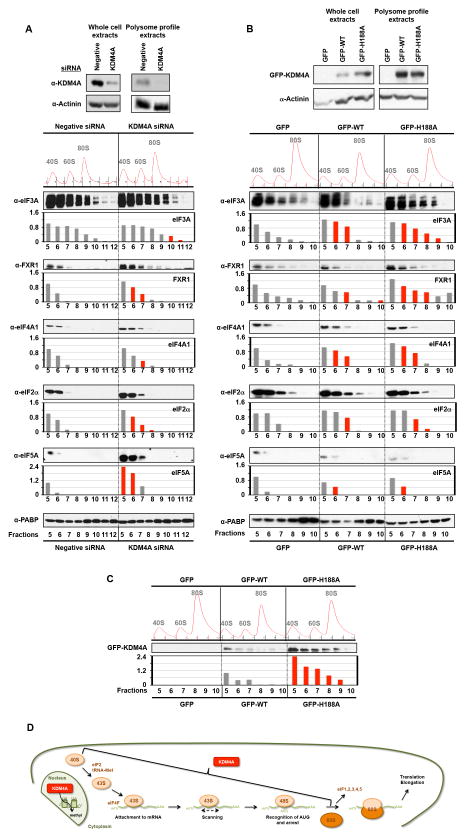
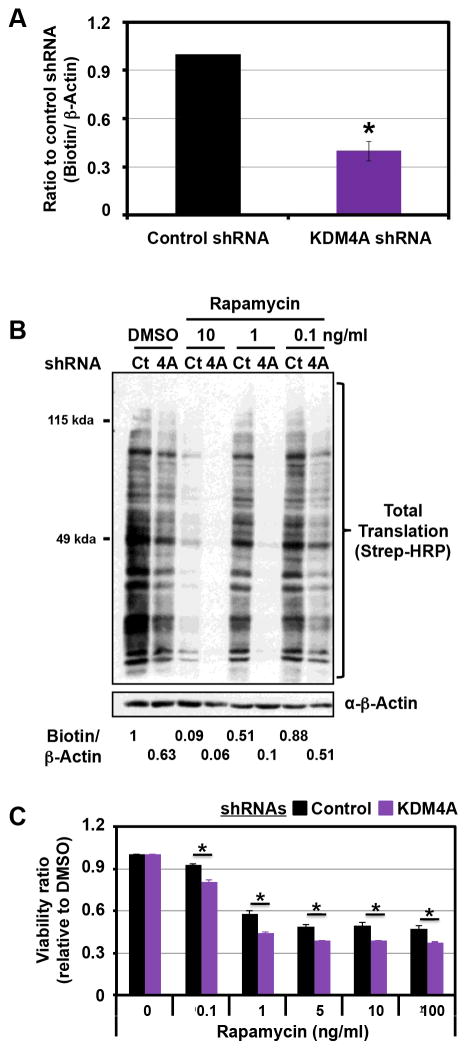
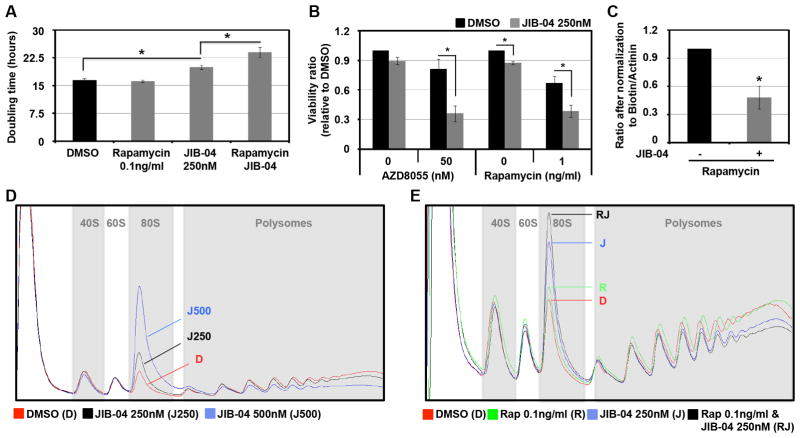
Comment in
-
From histones to ribosomes: a chromatin regulator tangoes with translation.Cancer Discov. 2015 Mar;5(3):228-30. doi: 10.1158/2159-8290.CD-15-0073. Cancer Discov. 2015. PMID: 25749972 Free PMC article.
References
-
- Su IH, Dobenecker MW, Dickinson E, Oser M, Basavaraj A, Marqueron R, et al. Polycomb group protein ezh2 controls actin polymerization and cell signaling. Cell. 2005;121:425–36. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
