

The kinetochore protein, CENPF, is mutated in human ciliopathy and microcephaly phenotypes
- PMID: 25564561
- PMCID: PMC4345935
- DOI: 10.1136/jmedgenet-2014-102691
The kinetochore protein, CENPF, is mutated in human ciliopathy and microcephaly phenotypes
Erratum in
-
Erratum: The kinetochore protein, CENPF, is mutated in human ciliopathy and microcephaly phenotypes.J Med Genet. 2016 Dec;53(12):865. doi: 10.1136/jmedgenet-2014-102691corr1. Epub 2016 Oct 13. J Med Genet. 2016. PMID: 27738186 Free PMC article. No abstract available.
Abstract
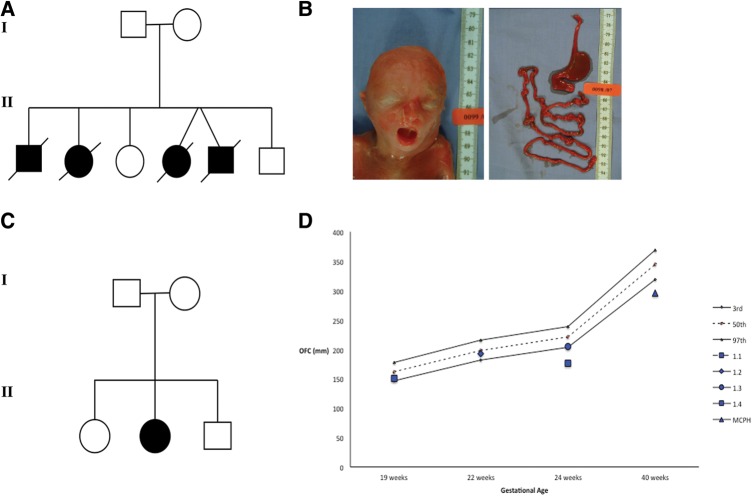
Background: Mutations in microtubule-regulating genes are associated with disorders of neuronal migration and microcephaly. Regulation of centriole length has been shown to underlie the pathogenesis of certain ciliopathy phenotypes. Using a next-generation sequencing approach, we identified mutations in a novel centriolar disease gene in a kindred with an embryonic lethal ciliopathy phenotype and in a patient with primary microcephaly.
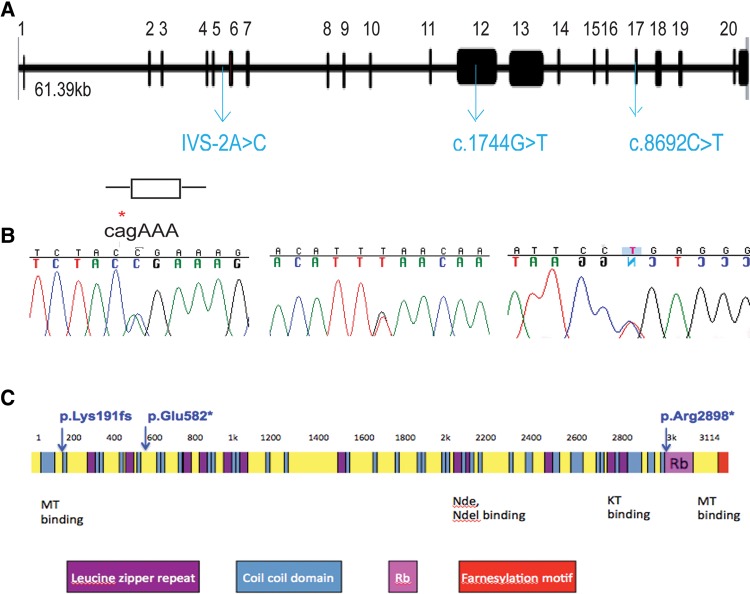
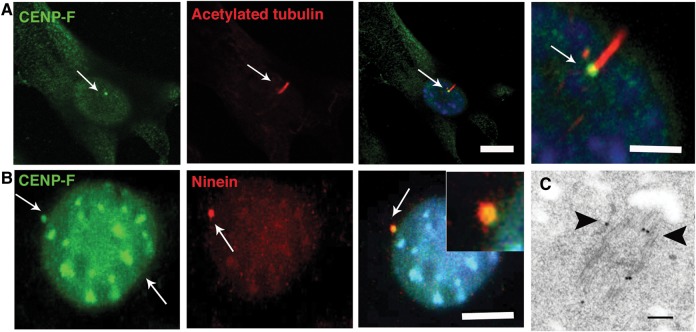
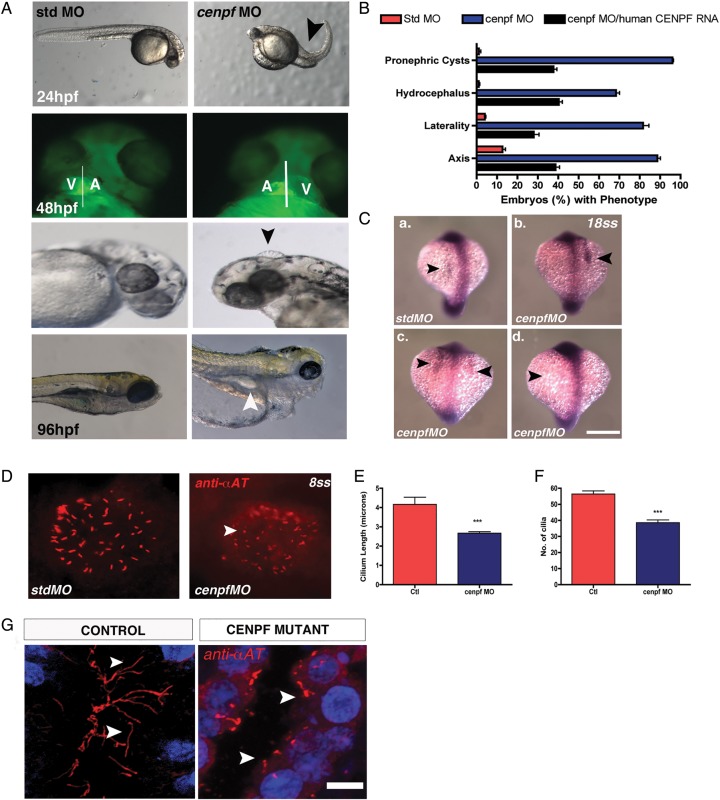
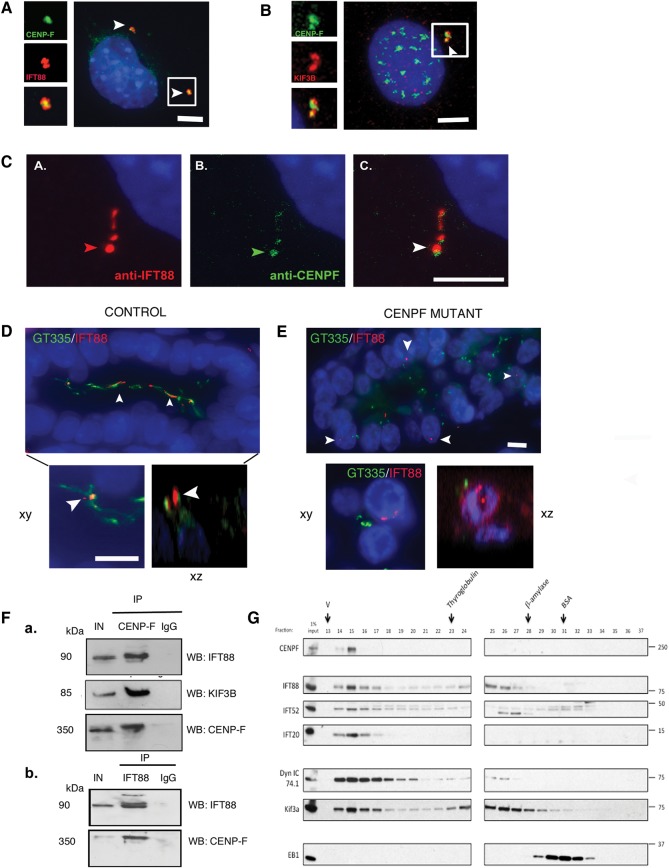
Methods and results: Whole exome sequencing data from a non-consanguineous Caucasian kindred exhibiting mid-gestation lethality and ciliopathic malformations revealed two novel non-synonymous variants in CENPF, a microtubule-regulating gene. All four affected fetuses showed segregation for two mutated alleles [IVS5-2A>C, predicted to abolish the consensus splice-acceptor site from exon 6; c.1744G>T, p.E582X]. In a second unrelated patient exhibiting microcephaly, we identified two CENPF mutations [c.1744G>T, p.E582X; c.8692 C>T, p.R2898X] by whole exome sequencing. We found that CENP-F colocalised with Ninein at the subdistal appendages of the mother centriole in mouse inner medullary collecting duct cells. Intraflagellar transport protein-88 (IFT-88) colocalised with CENP-F along the ciliary axonemes of renal epithelial cells in age-matched control human fetuses but did not in truncated cilia of mutant CENPF kidneys. Pairwise co-immunoprecipitation assays of mitotic and serum-starved HEKT293 cells confirmed that IFT88 precipitates with endogenous CENP-F.
Conclusions: Our data identify CENPF as a new centriolar disease gene implicated in severe human ciliopathy and microcephaly related phenotypes. CENP-F has a novel putative function in ciliogenesis and cortical neurogenesis.
Keywords: CENPF; Ciliopathy; Clinical genetics; Microcephaly; Molecular genetics.
Published by the BMJ Publishing Group Limited. For permission to use (where not already granted under a licence) please go to http://group.bmj.com/group/rights-licensing/permissions.
Figures
References
-
- Bornens M. The centrosome in cells and organisms. Science 2012;335:422–6. - PubMed
-
- Reiner O, Carrozzo R, Shen Y, Wehnert M, Faustinella F, Dobyns WB, Caskey CT, Ledbetter DH. Isolation of a Miller-Dieker lissencephaly gene containing G protein beta-subunit-like repeats. Nature 1993;364:717–21. - PubMed
-
- Rauch A, Thiel CT, Schindler D, Wick U, Crow YJ, Ekici AB, van Essen AJ, Goecke TO, Al-Gazali L, Chrzanowska KH, Zweier C, Brunner HG, Becker K, Curry CJ, Dallapiccola B, Devriendt K, Dörfler A, Kinning E, Megarbane A, Meinecke P, Semple RK, Spranger S, Toutain A, Trembath RC, Voss E, Wilson L, Hennekam R, de Zegher F, Dörr HG, Reis A. Mutations in the pericentrin (PCNT) gene cause primordial dwarfism. Science 2008;319:816–19. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases