

The role of alterations in mitochondrial dynamics and PGC-1α over-expression in fast muscle atrophy following hindlimb unloading
- PMID: 25565653
- PMCID: PMC4405755
- DOI: 10.1113/jphysiol.2014.286740
The role of alterations in mitochondrial dynamics and PGC-1α over-expression in fast muscle atrophy following hindlimb unloading
Abstract
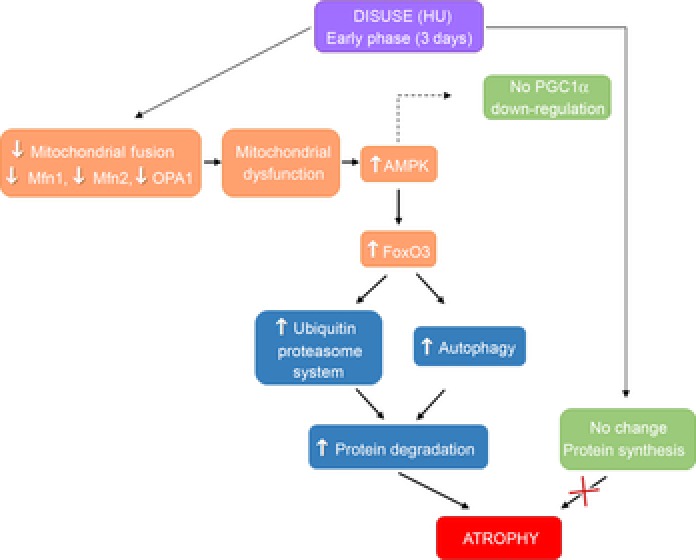
Key points: Skeletal muscle atrophy occurs as a result of disuse. Although several studies have established that a decrease in protein synthesis and increase in protein degradation lead to muscle atrophy, little is known about the triggers underlying such processes. A growing body of evidence challenges oxidative stress as a trigger of disuse atrophy; furthermore, it is also becoming evident that mitochondrial dysfunction may play a causative role in determining muscle atrophy. Mitochondrial fusion and fission have emerged as important processes that govern mitochondrial function and PGC-1α may regulate fusion/fission events. Although most studies on mice have focused on the anti-gravitary slow soleus muscle as it is preferentially affected by disuse atrophy, several fast muscles (including gastrocnemius) go through a significant loss of mass following unloading. Here we found that in fast muscles an early down-regulation of pro-fusion proteins, through concomitant AMP-activated protein kinase (AMPK) activation, can activate catabolic systems, and ultimately cause muscle mass loss in disuse. Elevated muscle PGC-1α completely preserves muscle mass by preventing the fall in pro-fusion protein expression, AMPK and catabolic system activation, suggesting that compounds inducing PGC-1α expression could be useful to treat and prevent muscle atrophy.
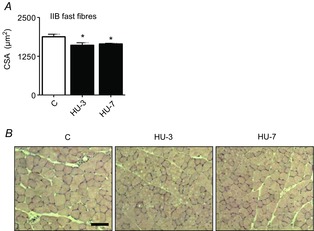
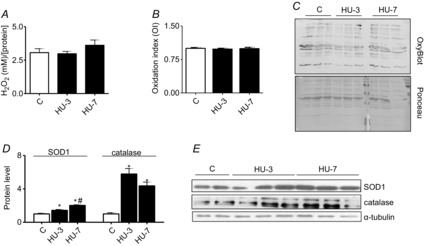
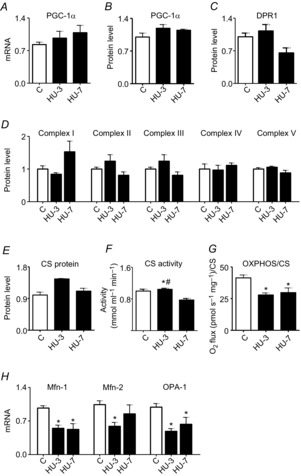
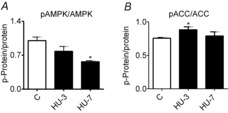
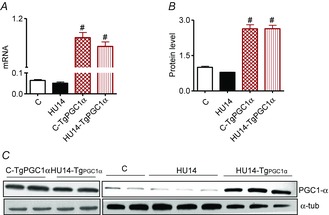
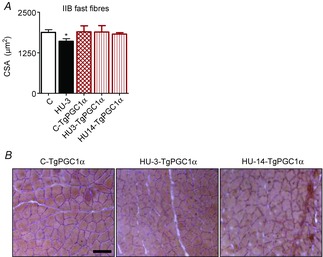
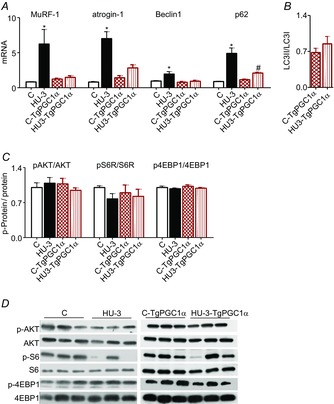
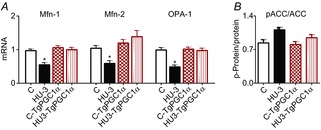
Abstract: The mechanisms triggering disuse muscle atrophy remain of debate. It is becoming evident that mitochondrial dysfunction may regulate pathways controlling muscle mass. We have recently shown that mitochondrial dysfunction plays a major role in disuse atrophy of soleus, a slow, oxidative muscle. Here we tested the hypothesis that hindlimb unloading-induced atrophy could be due to mitochondrial dysfunction in fast muscles too, notwithstanding their much lower mitochondrial content. Gastrocnemius displayed atrophy following both 3 and 7 days of unloading. SOD1 and catalase up-regulation, no H2 O2 accumulation and no increase of protein carbonylation suggest the antioxidant defence system efficiently reacted to redox imbalance in the early phases of disuse. A defective mitochondrial fusion (Mfn1, Mfn2 and OPA1 down-regulation) occurred together with an impairment of OXPHOS capacity. Furthermore, at 3 days of unloading higher acetyl-CoA carboxylase (ACC) phosphorylation was found, suggesting AMP-activated protein kinase (AMPK) pathway activation. To test the role of mitochondrial alterations we used Tg-mice overexpressing PGC-1α because of the known effect of PGC-1α on stimulation of Mfn2 expression. PGC-α overexpression was sufficient to prevent (i) the decrease of pro-fusion proteins (Mfn1, Mfn2 and OPA1), (ii) activation of the AMPK pathway, (iii) the inducible expression of MuRF1 and atrogin1 and of authopagic factors, and (iv) any muscle mass loss in response to disuse. As the effects of increased PGC-1α activity were sustained throughout disuse, compounds inducing PGC-1α expression could be useful to treat and prevent muscle atrophy also in fast muscles.
© 2015 The Authors. The Journal of Physiology © 2015 The Physiological Society.
Figures
Comment in
-
Regulation of muscle mass: a new role for mitochondria?J Physiol. 2015 Apr 15;593(8):1761-2. doi: 10.1113/JP270216. J Physiol. 2015. PMID: 25656316 Free PMC article. No abstract available.
-
Towards a better understanding of the role played by mitochondrial dynamics and morphology in skeletal muscle atrophy.J Physiol. 2015 Jul 15;593(14):2993-4. doi: 10.1113/JP270736. J Physiol. 2015. PMID: 26173825 Free PMC article. No abstract available.
References
-
- Berchtold MW, Brinkmeier H. Müntener M. Calcium ion in skeletal muscle: its crucial role for muscle function, plasticity, and disease. Physiol Rev. 2000;80:1215–1265. - PubMed
-
- Bodine SC, Latres E, Baumhueter S, Lai VK, Nunez L, Clarke BA, Poueymirou WT, Panaro FJ, Na E, Dharmarajan K, Pan ZQ, Valenzuela DM, DeChiara TM, Stitt TN, Yancopoulos GD. Glass DJ. Identification of ubiquitin ligases required for skeletal muscle atrophy. Science. 2001a;294:1704–1708. - PubMed
-
- Bodine SC, Stitt TN, Gonzalez M, Kline WO, Stover GL, Bauerlein R, Zlotchenko E, Scrimgeour A, Lawrence JC, Glass DJ. Yancopoulos GD. Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nat Cell Biol. 2001b;3:1014–1019. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous