

Irinotecan treatment and senescence failure promote the emergence of more transformed and invasive cells that depend on anti-apoptotic Mcl-1
- PMID: 25565667
- PMCID: PMC4381604
- DOI: 10.18632/oncotarget.2774
Irinotecan treatment and senescence failure promote the emergence of more transformed and invasive cells that depend on anti-apoptotic Mcl-1
Abstract
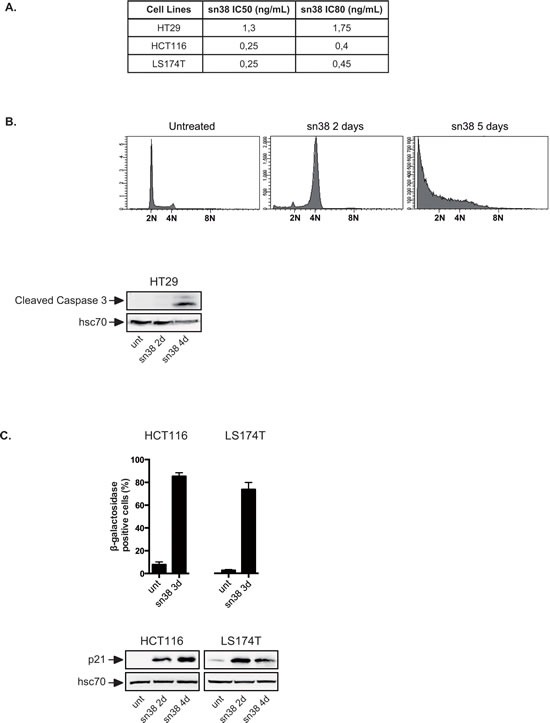
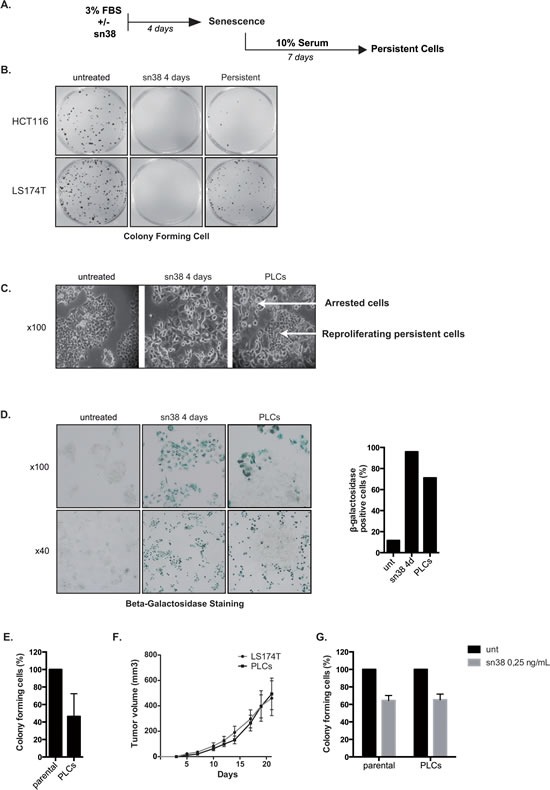
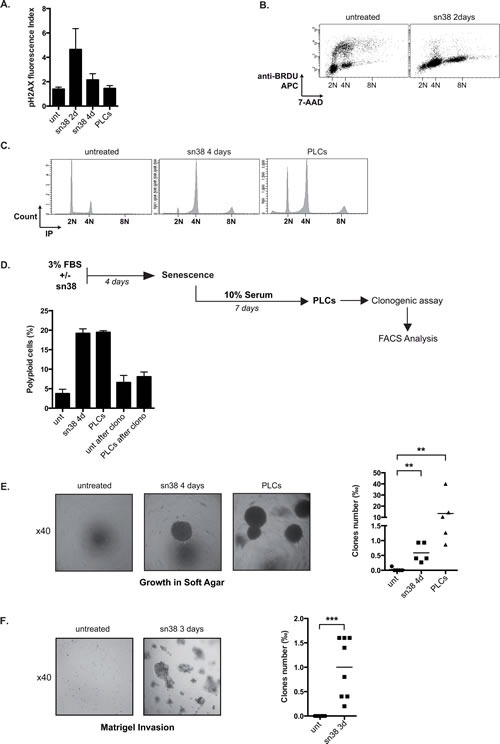
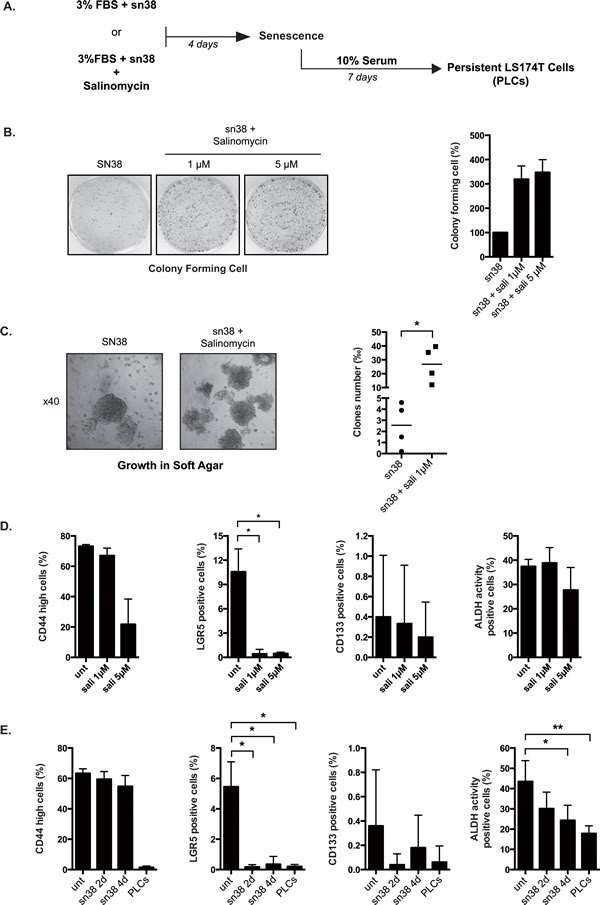
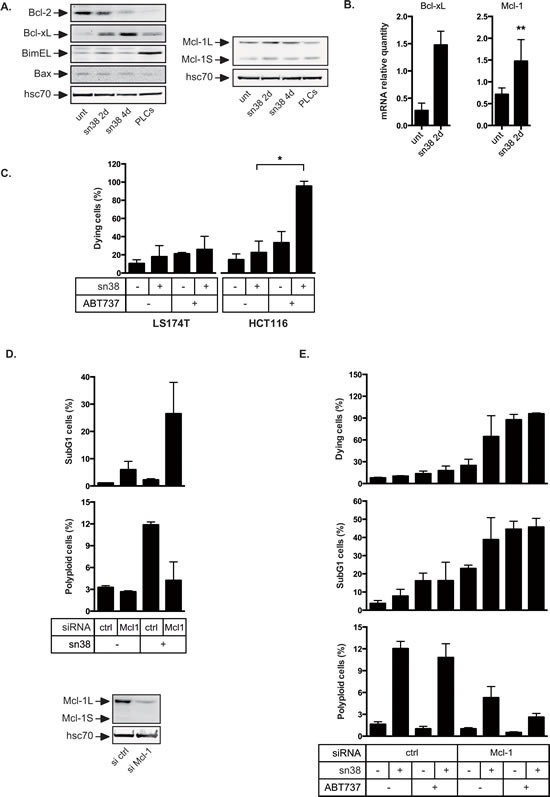
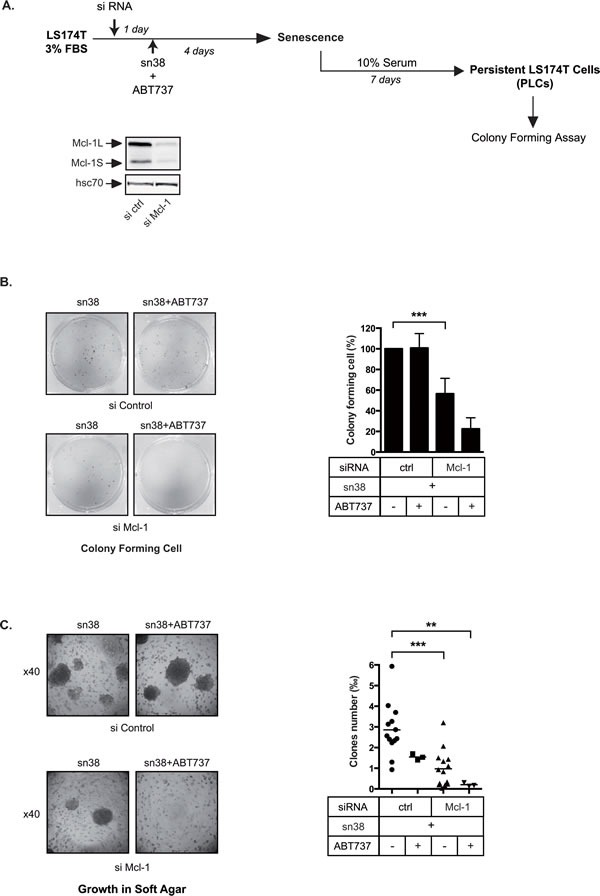
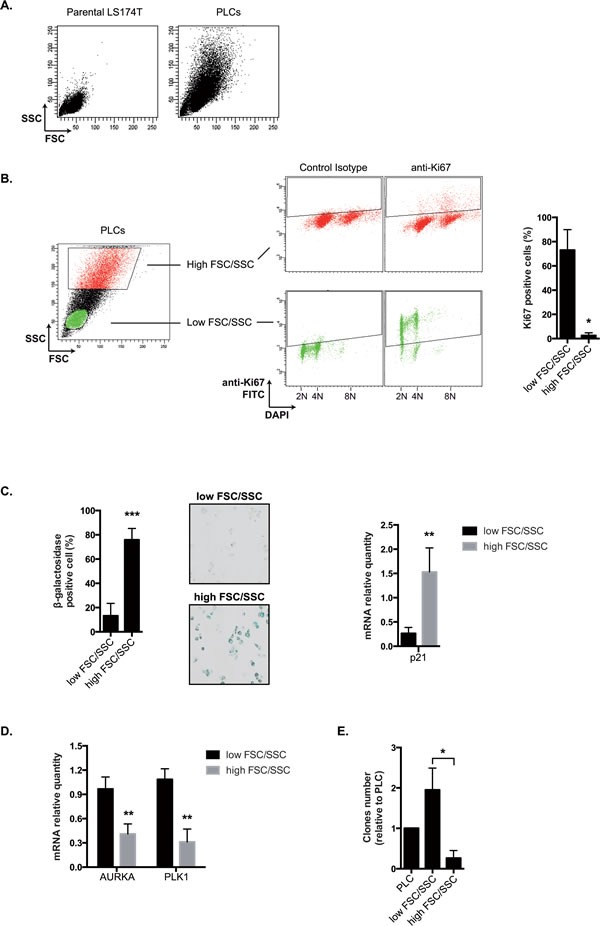
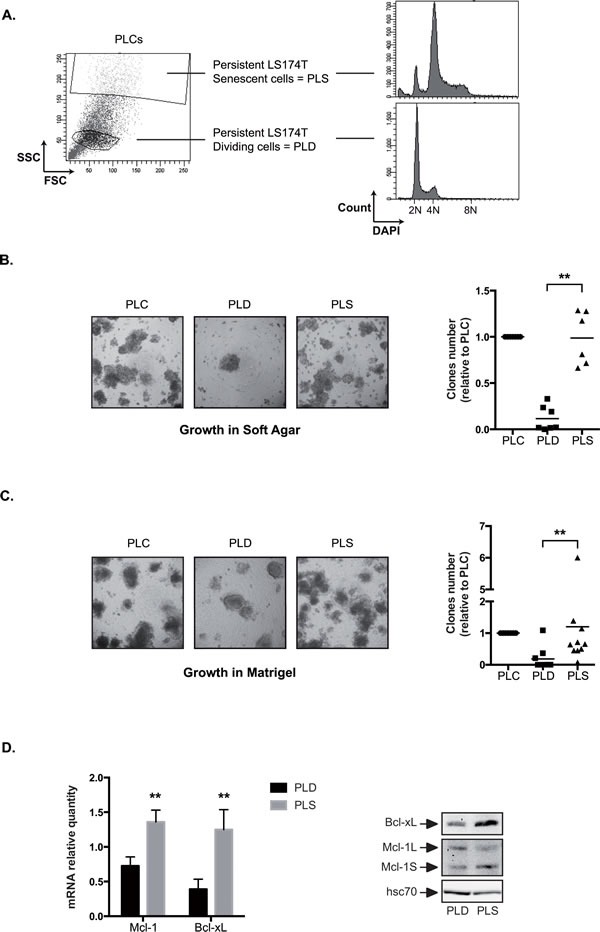
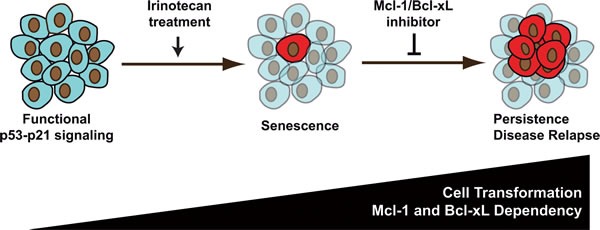
Induction of senescence by chemotherapy was initially characterized as a suppressive response that prevents tumor cell proliferation. However, in response to treatment, it is not really known how cells can survive senescence and how irreversible this pathway is. In this study, we analyzed cell escape in response to irinotecan, a first line treatment used in colorectal cancer that induced senescence. We detected subpopulations of cells that adapted to chemotherapy and resumed proliferation. Survival led to the emergence of more transformed cells that induced tumor formation in mice and grew in low adhesion conditions. A significant amount of viable polyploid cells was also generated following irinotecan failure. Markers such as lgr5, CD44, CD133 and ALDH were downregulated in persistent clones, indicating that survival was not associated with an increase in cancer initiating cells. Importantly, malignant cells which resisted senescence relied on survival pathways induced by Mcl-1 signaling and to a lesser extent by Bcl-xL. Depletion of Mcl-1 increased irinotecan efficiency, induced the death of polyploid cells, prevented cell emergence and inhibited growth in low-adhesion conditions. We therefore propose that Mcl-1 targeting should be considered in the future to reduce senescence escape and to improve the treatment of irinotecan-refractory colorectal cancers.
Figures
Comment in
-
Mcl-1 and tumor cell persistence.Oncotarget. 2015 Jan 1;6(1):5-6. doi: 10.18632/oncotarget.3035. Oncotarget. 2015. PMID: 25575820 Free PMC article. No abstract available.
-
Senescence versus apoptosis in chemotherapy.Oncotarget. 2015 Mar 10;6(7):4551-2. doi: 10.18632/oncotarget.3114. Oncotarget. 2015. PMID: 25732846 Free PMC article. No abstract available.
References
-
- Narita M, Nunez S, Heard E, Narita M, Lin AW, Hearn SA, Spector DL, Hannon GJ, Lowe SW. Rb-mediated heterochromatin formation and silencing of E2F target genes during cellular senescence. Cell. 2003;113(6):703–716. - PubMed
-
- Roninson IB. Tumor cell senescence in cancer treatment. Cancer Res. 2003;63(11):2705–2715. - PubMed
-
- Schmitt CA, Fridman JS, Yang M, Lee S, Baranov E, Hoffman RM, Lowe SW. A senescence program controlled by p53 and p16INK4a contributes to the outcome of cancer therapy. Cell. 2002;109(3):335–346. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
