

The heterogeneity in GABAA receptor-mediated IPSC kinetics reflects heterogeneity of subunit composition among inhibitory and excitatory interneurons in spinal lamina II
- PMID: 25565959
- PMCID: PMC4263102
- DOI: 10.3389/fncel.2014.00424
The heterogeneity in GABAA receptor-mediated IPSC kinetics reflects heterogeneity of subunit composition among inhibitory and excitatory interneurons in spinal lamina II
Abstract
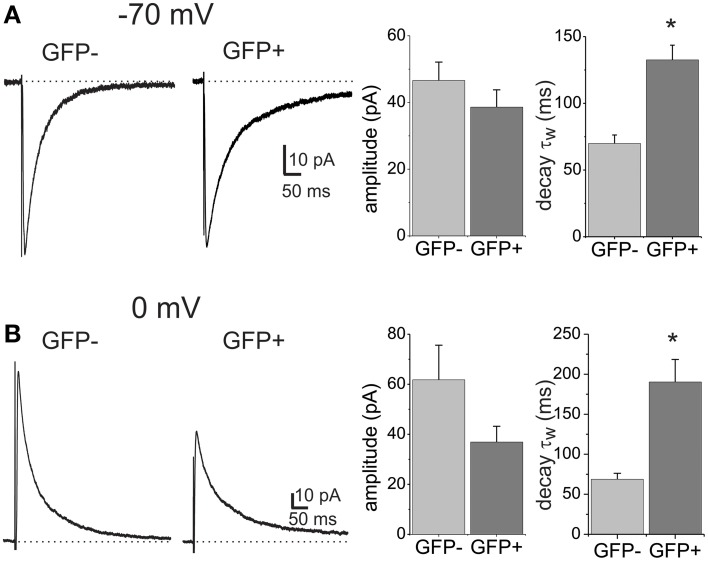
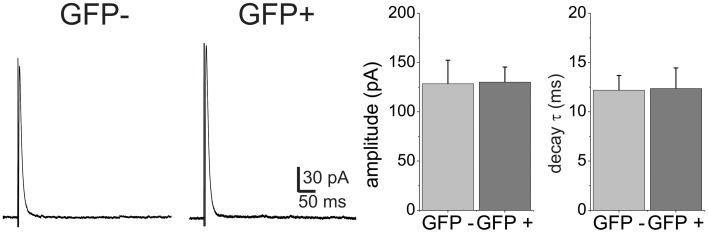
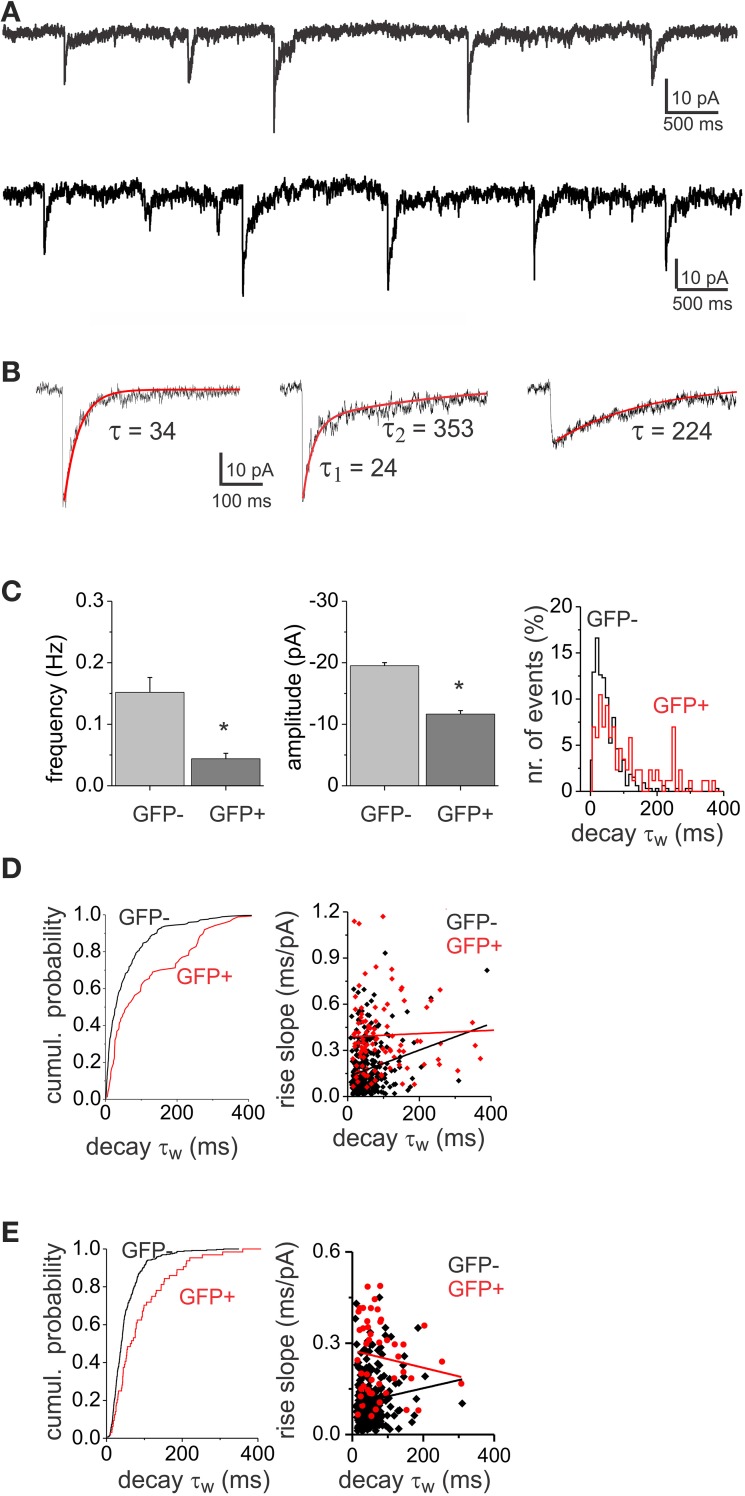
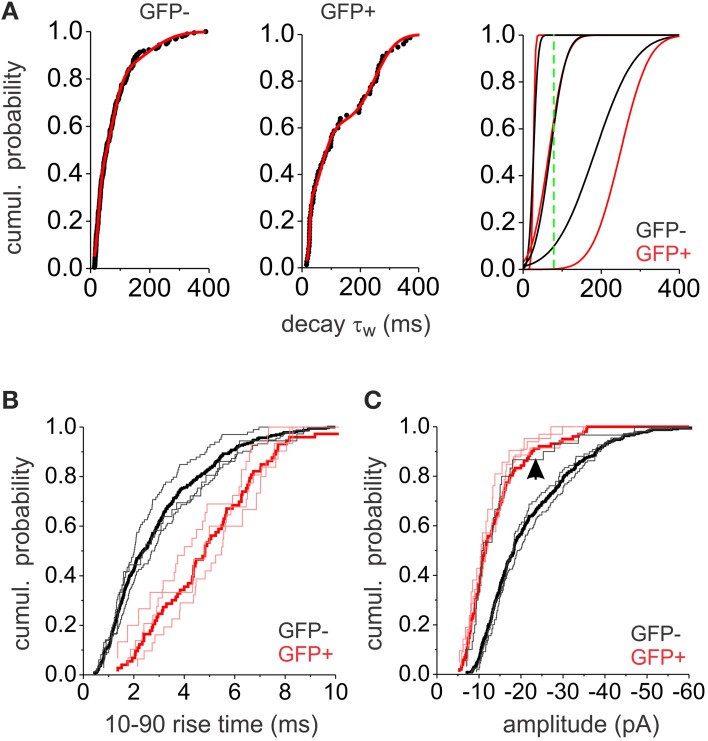
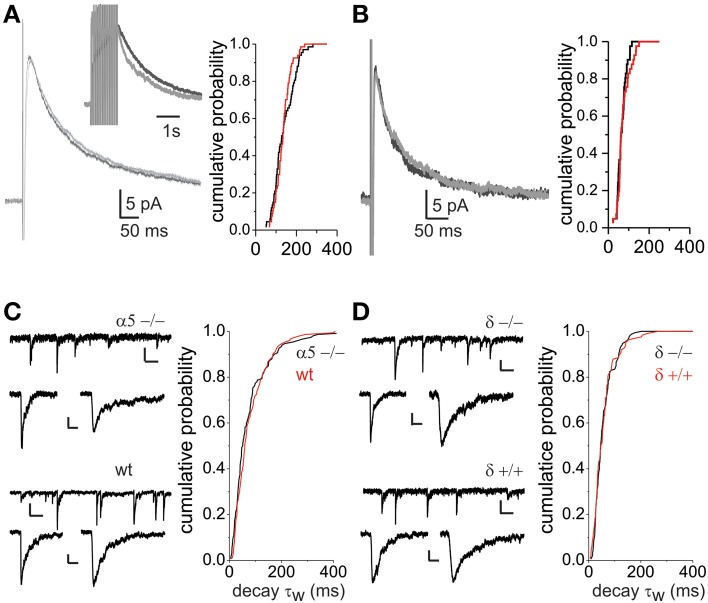
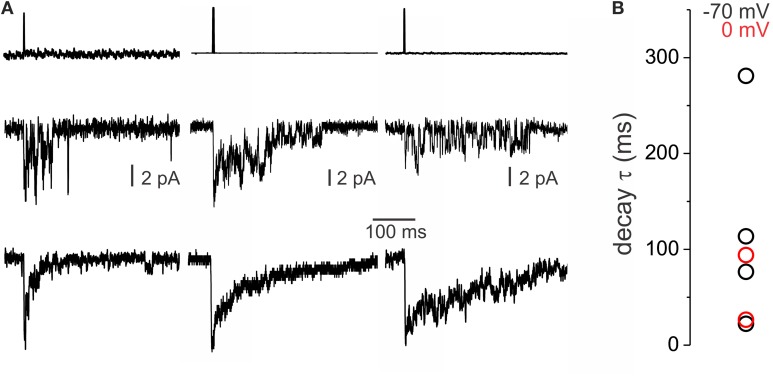
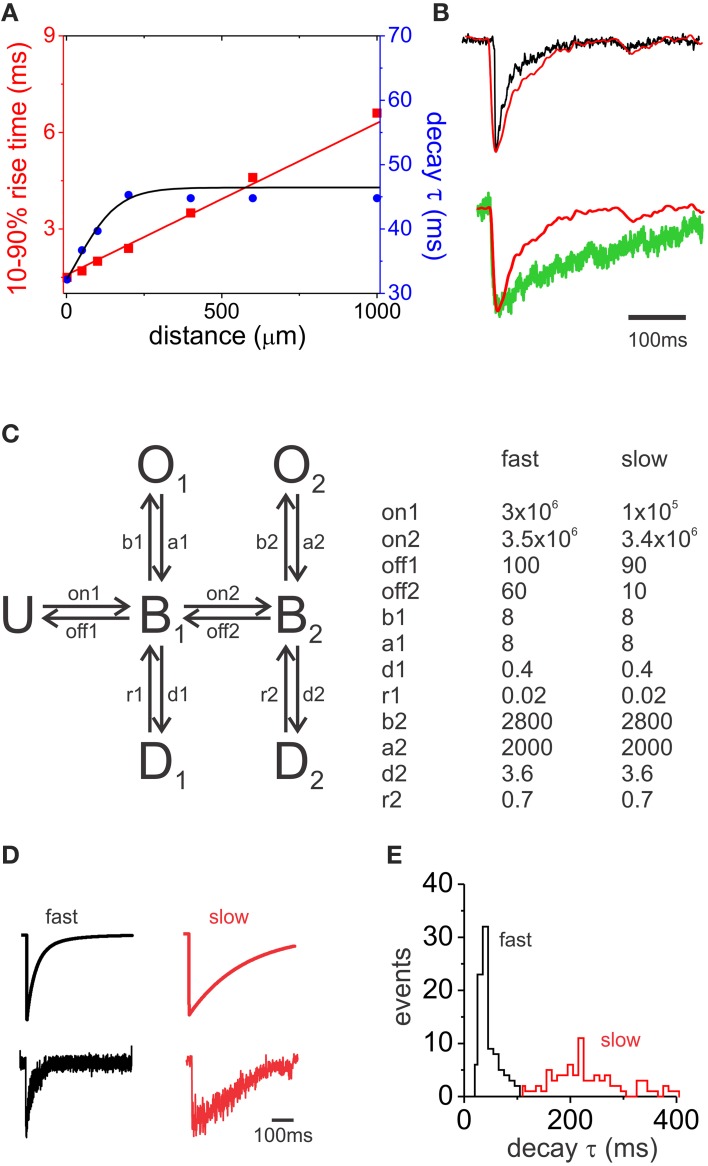
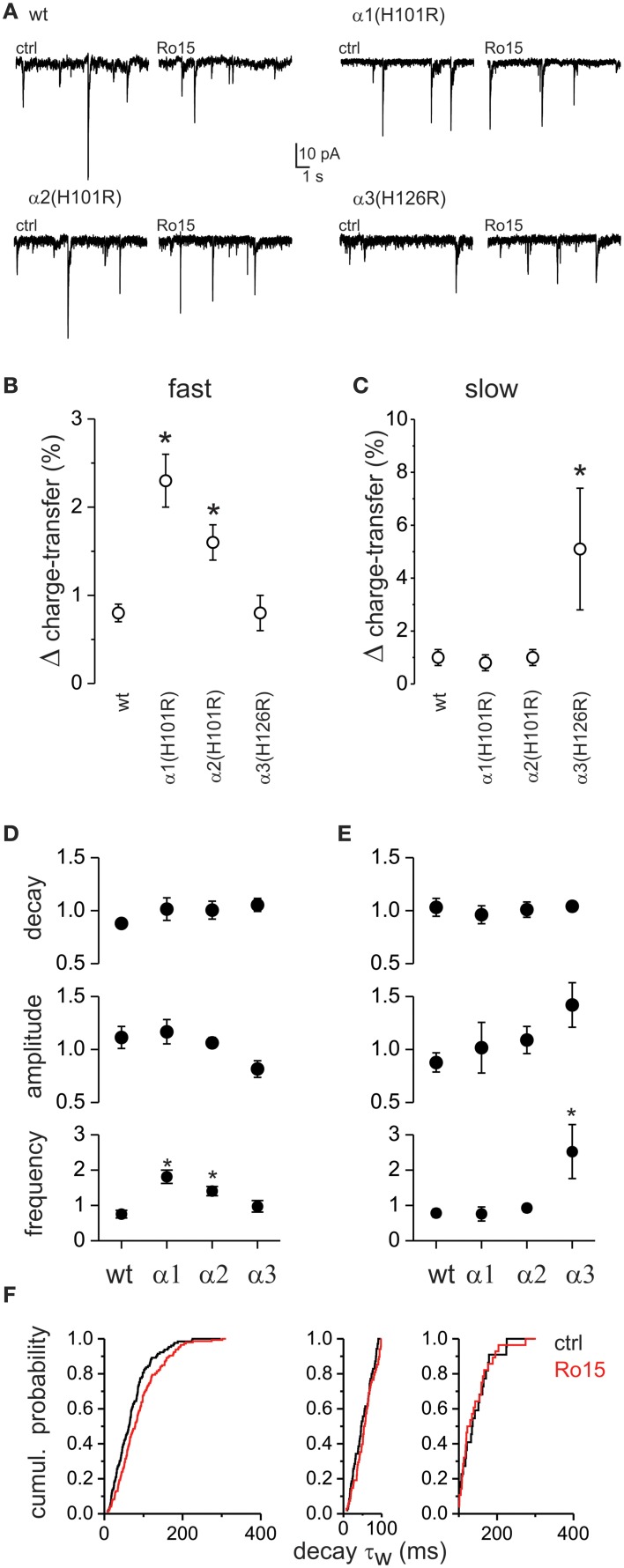
GABAergic inhibition displays rich functional diversity throughout the CNS, which arises from variations in the nature of inputs, subunit composition, subcellular localization of receptors and synapse geometry, or reuptake mechanisms. In the spinal dorsal horn (SDH), GABAA and glycine receptors play a major role in the control of excitability and accuracy of nociceptive processing. Identifying which components shape the properties of the inhibitory synapses in different cell types is necessary to understand how nociceptive information is integrated. To address this, we used transgenic mice where inhibitory interneurons express GAD65-EGFP. We found that GABAA, but not glycine receptor-mediated evoked IPSCs displayed slower kinetics in EGFP+ vs. EGFP- interneurons. GABAA miniature IPSC decay kinetics showed a large variability in both populations, however the distribution of decays differed between EGFP+ and EGFP- interneurons. The range of mIPSC decay kinetics observed was replicated in experiments using rapid application of GABA on outside-out patches taken from SDH neurons in slices. Furthermore, GABAA decay kinetics were not affected by uptake blockers and were not different in mice lacking δ or α5 subunits, indicating that intrinsic channel properties likely underlie the heterogeneity. To identify whether other α subunits shape the various kinetic properties observed we took advantage of knock-in mice carrying point mutations in either the α1, α2, or α3 subunits rendering Ro 15-4513 a selective agonist at the benzodiazepine modulatory site. We found that α1 and α2 subunit underlie the fast decaying component of IPSCs while the slow component is determined by the α3 subunit. The differential distribution of GABAA subunits at inhibitory synapses thus sculpts the heterogeneity of the SDH inhibitory circuitry. This diversity of inhibitory elements can be harnessed to selectively modulate different components of the spinal nociceptive circuitry for therapeutic interventions.
Keywords: GABAA receptors; IPSCs; decay kinetics; spinal dorsal horn; subunit composition.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials