

Differential responses to acute administration of a new 5-HT7-R agonist as a function of adolescent pre-treatment: phMRI and immuno-histochemical study
- PMID: 25565998
- PMCID: PMC4267273
- DOI: 10.3389/fnbeh.2014.00427
Differential responses to acute administration of a new 5-HT7-R agonist as a function of adolescent pre-treatment: phMRI and immuno-histochemical study
Abstract
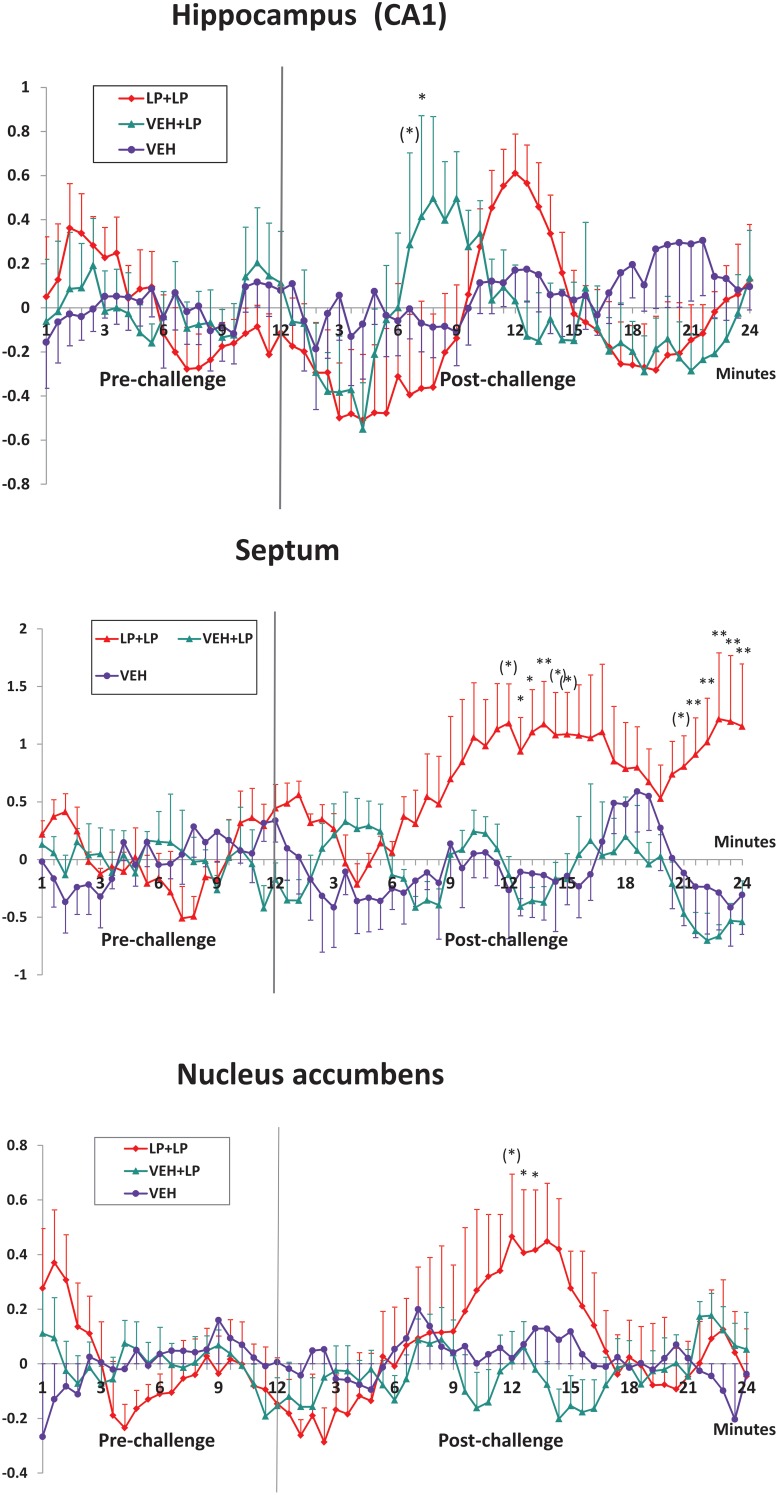
LP-211 is a new, selective agonist of serotonin (5-hydroxytryptamine, 5-HT) receptor 7 (5-HT7-R), which is part of a neuro-transmission system with a proposed role in neural plasticity and in mood, cognitive and sleep regulation. Adolescent subchronic LP-211 treatment produces some persisting changes in rats' forebrain structural and functional parameters. Here, using pharmacological MRI (phMRI), we investigated the effect of acute administration with LP-211 (10 mg/kg i.p.), or vehicle, to adult rats previously exposed to the same drug (0.25 mg/kg/day for 5 days), or vehicle, during adolescence (44-48 post-natal days); histology and immuno-histochemistry were performed ex vivo to evaluate neuro-anatomical and physiological long-term adaptation to pharmacological pre-treatment. The phMRI signal reveals forebrain areas (i.e., hippocampus, orbital prefrontal cortex), activated in response to LP-211 challenge independently of adolescent pre-treatment. In septum and nucleus accumbens, sensitized activation was found in adolescent pre-treated rats but not in vehicle-exposed controls. Immuno-histochemical analyses showed marked differences in septum as long-term consequence of the adolescent pre-treatment: increased level of 5-HT7-R, increased number of 5-HT7-R positive cells, and enhanced astrocyte activation. For nucleus accumbens, immuno-histochemical analyses did not reveal any difference between adolescent pre-treated rats and vehicle-exposed controls. In conclusion, subchronic LP-211 administration during adolescence is able to induce persistent physiological changes in the septal 5-HT7-R expression and astrocyte response that can still be observed in adulthood. Data shed new insights into roles of 5-HT7-R for normal and pathologic behavioral regulations.
Keywords: LP-211; Ph-MRI; hippocampus; limbic/cortical loop; nucleus accumbens; septum.
Figures




References
-
- Abdi H., Williams L. J. (2010). Tukey's honestly significant difference (HSD) test. Encyclopedia Res. Des. 1, 1–5 10.4135/9781412961288.n181 - DOI
-
- Bonaventure P., Nepomuceno D., Hein L., Sutcliffe J. G., Lovenberg T., Hedlund P. B. (2004). Radioligand binding analysis of knockout mice reveals 5-hydroxytryptamine(7) receptor distribution and uncovers 8-hydroxy-2-(di-n-propylamino)tetralin interaction with alpha(2) adrenergic receptors. Neuroscience 124, 901–911. 10.1016/j.neuroscience.2004.01.014 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources