

Small tRNA-derived RNAs are increased and more abundant than microRNAs in chronic hepatitis B and C
- PMID: 25567797
- PMCID: PMC4286764
- DOI: 10.1038/srep07675
Small tRNA-derived RNAs are increased and more abundant than microRNAs in chronic hepatitis B and C
Abstract
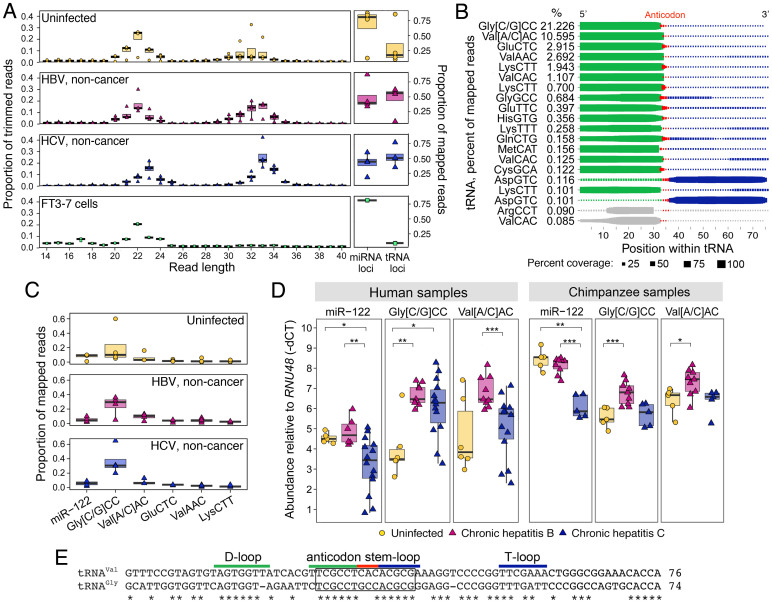
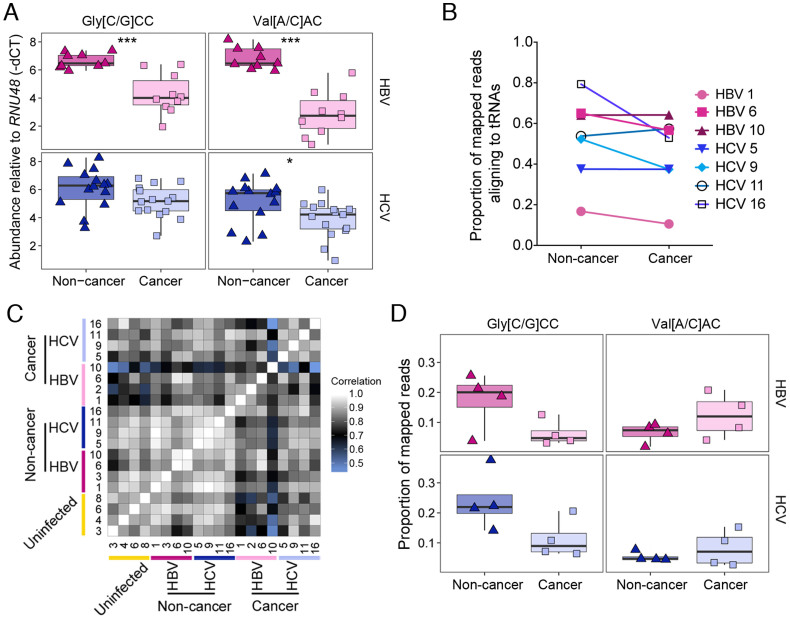
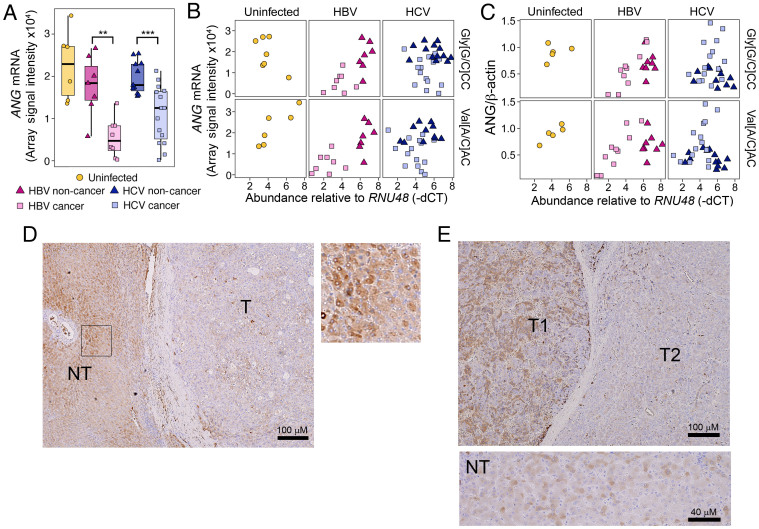
Persistent infections with hepatitis B virus (HBV) or hepatitis C virus (HCV) account for the majority of cases of hepatic cirrhosis and hepatocellular carcinoma (HCC) worldwide. Small, non-coding RNAs play important roles in virus-host interactions. We used high throughput sequencing to conduct an unbiased profiling of small (14-40 nts) RNAs in liver from Japanese subjects with advanced hepatitis B or C and hepatocellular carcinoma (HCC). Small RNAs derived from tRNAs, specifically 30-35 nucleotide-long 5' tRNA-halves (5' tRHs), were abundant in non-malignant liver and significantly increased in humans and chimpanzees with chronic viral hepatitis. 5' tRH abundance exceeded microRNA abundance in most infected non-cancerous tissues. In contrast, in matched cancer tissue, 5' tRH abundance was reduced, and relative abundance of individual 5' tRHs was altered. In hepatitis B-associated HCC, 5' tRH abundance correlated with expression of the tRNA-cleaving ribonuclease, angiogenin. These results demonstrate that tRHs are the most abundant small RNAs in chronically infected liver and that their abundance is altered in liver cancer.
Figures
References
-
- Arzumanyan A., Reis H. M. & Feitelson M. A. Pathogenic mechanisms in HBV- and HCV-associated hepatocellular carcinoma. Nat Rev Cancer 13, 123–135 (2013). - PubMed
-
- Perz J. F., Armstrong G. L., Farrington L. A., Hutin Y. J. & Bell B. P. The contributions of hepatitis B virus and hepatitis C virus infections to cirrhosis and primary liver cancer worldwide. J Hepatol 45, 529–538 (2006). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- C06 RR12087/RR/NCRR NIH HHS/United States
- C06 RR016228/RR/NCRR NIH HHS/United States
- OD P51 OD011133/OD/NIH HHS/United States
- R01 AI095690/AI/NIAID NIH HHS/United States
- T32 AI007419/AI/NIAID NIH HHS/United States
- T32 GM067553/GM/NIGMS NIH HHS/United States
- T32-GM067553/GM/NIGMS NIH HHS/United States
- R01 CA164029/CA/NCI NIH HHS/United States
- R00-DK091318/DK/NIDDK NIH HHS/United States
- R01-AI095690/AI/NIAID NIH HHS/United States
- T32-AI007419/AI/NIAID NIH HHS/United States
- R01-CA164029/CA/NCI NIH HHS/United States
- P51 OD011133/OD/NIH HHS/United States
- C06 RR012087/RR/NCRR NIH HHS/United States
- R00 DK091318/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
