

Nucleoid occlusion protein Noc recruits DNA to the bacterial cell membrane
- PMID: 25568309
- PMCID: PMC4331003
- DOI: 10.15252/embj.201490177
Nucleoid occlusion protein Noc recruits DNA to the bacterial cell membrane
Abstract
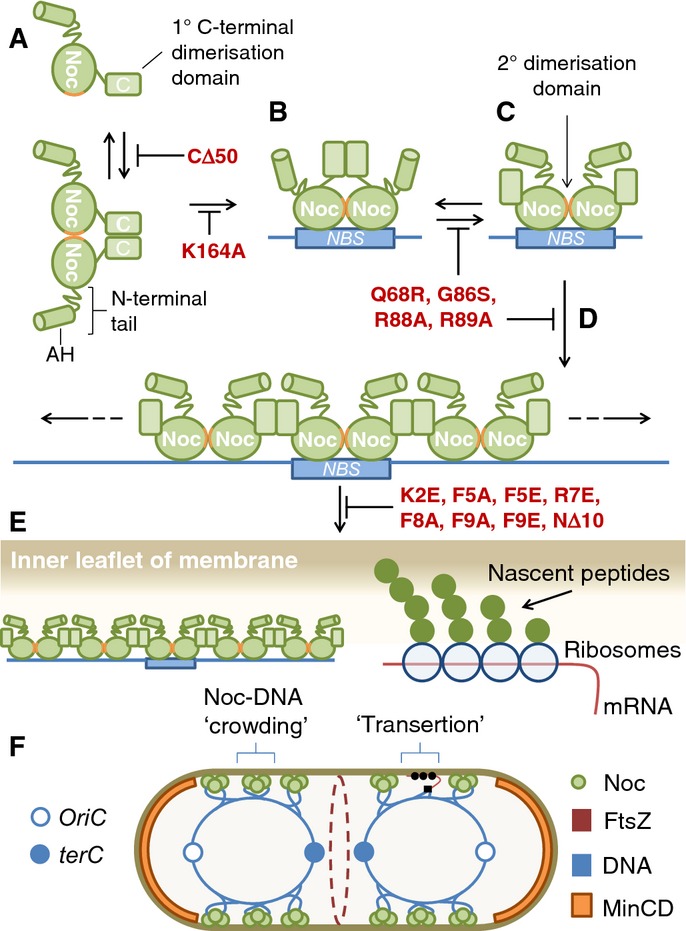
To proliferate efficiently, cells must co-ordinate division with chromosome segregation. In Bacillus subtilis, the nucleoid occlusion protein Noc binds to specific DNA sequences (NBSs) scattered around the chromosome and helps to protect genomic integrity by coupling the initiation of division to the progression of chromosome replication and segregation. However, how it inhibits division has remained unclear. Here, we demonstrate that Noc associates with the cell membrane via an N-terminal amphipathic helix, which is necessary for function. Importantly, the membrane-binding affinity of this helix is weak and requires the assembly of nucleoprotein complexes, thus establishing a mechanism for DNA-dependent activation of Noc. Furthermore, division inhibition by Noc requires recruitment of NBS DNA to the cell membrane and is dependent on its ability to bind DNA and membrane simultaneously. Indeed, Noc production in a heterologous system is sufficient for recruitment of chromosomal DNA to the membrane. Our results suggest a simple model in which the formation of large membrane-associated nucleoprotein complexes physically occludes assembly of the division machinery.
Keywords: Bacillus subtilis; FtsZ; Noc; ParB; nucleoid occlusion.
© 2015 The Authors. Published under the terms of the CC BY 4.0 license.
Figures
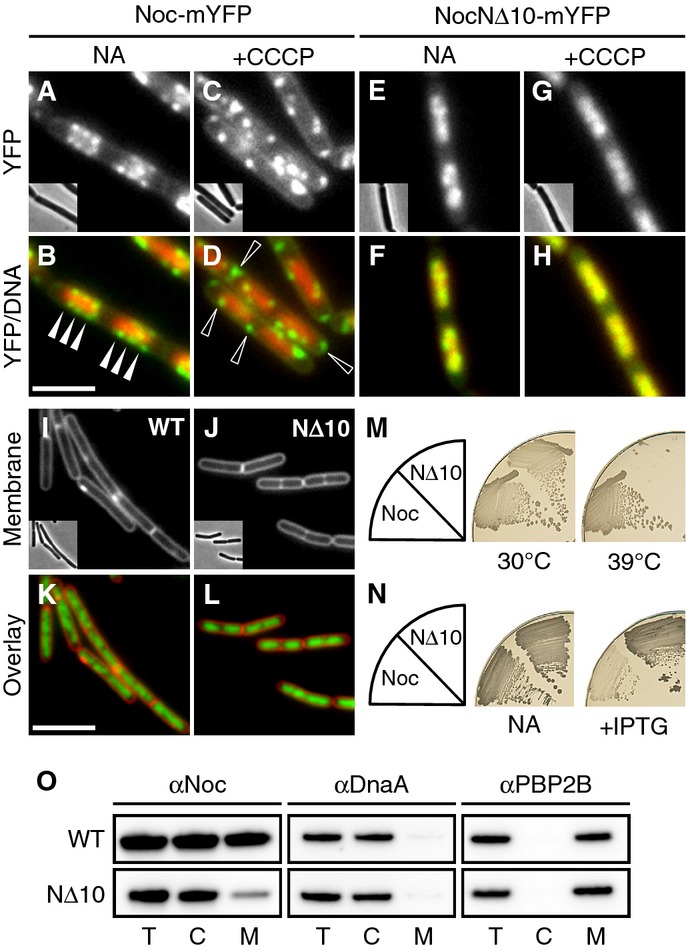
Effect of CCCP on the localisation of Noc and NocNΔ10. Cellular localisation of Noc-mYFP (DWA206) and NocNΔ10-mYFP (DWA382) either with no additions (NA) or after CCCP treatment (5 min; 100 μM), as indicated. Scale bar, 2.5 μm.
Effect of Noc overproduction on cell division. Exponentially growing cultures of DWA119 (Δnoc,Pspac(hy)-noc) and DWA282 (Δnoc,Pspac(hy)-nocNΔ10) were examined after growth for 1 h with 1 mM IPTG. Cell membranes and DNA were stained with FM5-95 and DAPI, respectively. Insets show corresponding phase contrast images. Scale bar, 5 μm.
Complementation of noc in a Δnoc ΔminCD background. Strains DWA564 (Δnoc, ΔminCD, Pxyl-noc-myfp) and 566 (Δnoc, ΔminCD, Pxyl-nocNΔ10-myfp) were streaked on nutrient agar (NA) plates in the presence of 0.5% w/v xylose and incubated at 30 and 39°C, as indicated.
Effect of Noc and NocNΔ10 overproduction on sporulation. Strains DWA119 (Δnoc,Pspac(hy)-noc) and DWA282 (Δnoc,Pspac(hy)-nocNΔ10) were streaked on NA plates in the absence and presence of 1 mM IPTG, as indicated, and photographed after 48 h at 37°C.
Western blot analysis of cellular fractions (T, total; C, cytosolic; M, membrane) of strains DWA119 and 282 expressing Noc or NocNΔ10, respectively. Proteins were detected using polyclonal antibodies against Noc, DnaA and PBP2B. Antibodies were used at a dilution of 1:10,000.
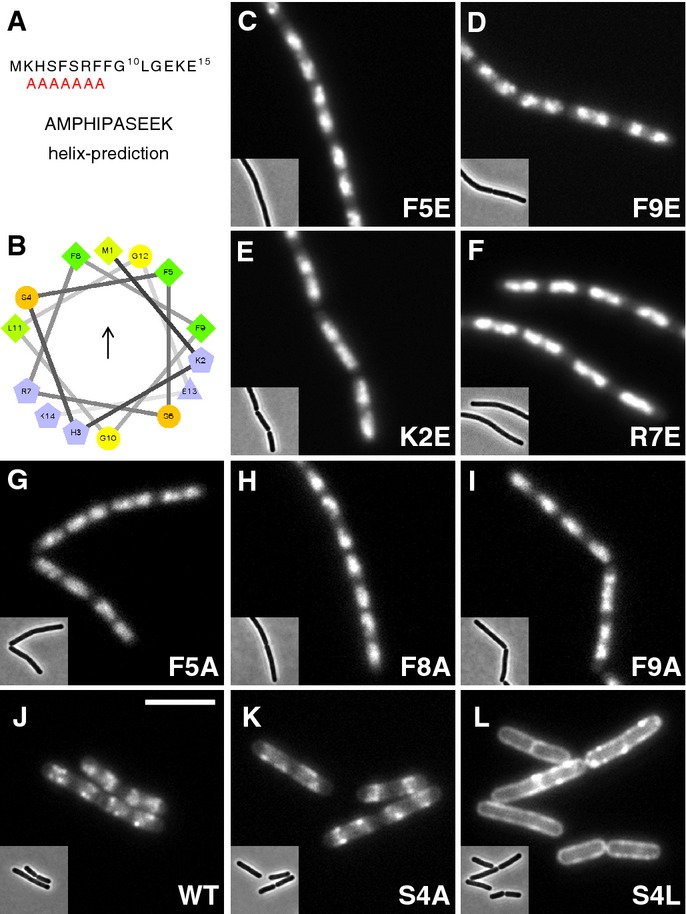
AMPHIPASEEK prediction result for Bacillus subtilis Noc. The red “A”s indicate a putative amphipathic helical region.
Helical wheel projection of the N-terminus (aa 1–14) showing the presence of hydrophobic (arrow) and polar faces. Residues are coloured according to their properties, greens, hydrophobic; blues, charged; orange, polar, uncharged; and yellow, glycine. The figure was prepared using the tool available at
http://rzlab.ucr.edu/scripts/wheel/wheel.cgi .Effects of N-terminal substitutions on Noc localisation, in strains: DWA211 (F5E), 318 (F9E), 316 (K2E), 212 (R7E), 322 (F5A), 323 (F8A), 325 (F9A), 206 (WT), 328 (S4A) and 329 (S4L), as indicated. Insets show the corresponding phase contrast images. Scale bar, 5 μm.

Helical wheel projection showing the amphipathic helix from hepatitis C virus protein NS4B. The hydrophobic face is indicated by an arrow. Colour scheme as in Fig2B.
Growth of strains DWA350 (Δnoc ΔminCD) and DWA307 (Δnoc ΔminCD,Pxyl-HCVAH-NocNΔ10) on nutrient agar plates at 30 and 48°C in the absence and presence of 0.5% w/v xylose, as indicated.
Cell morphology of strains DWA350 and DWA307 following growth in LB at 30°C (C and D) and at 42°C in LB + 0.5% w/v xylose (E and F). Arrowheads indicate minicells. Cell membranes were stained with FM5–95. Insets show the corresponding phase contrast images. Scale bar, 5 μm.
Cellular localisation of HCVAH-NocNΔ10-YFP (G) in strain DWA193 (Δnoc,Pxyl-HCVAH-nocNΔ10-yfp) and overlay showing DAPI-stained DNA (H). The strain was grown at 30°C in CH medium. Inset shows the corresponding phase contrast image. Scale bar, 5 μm.
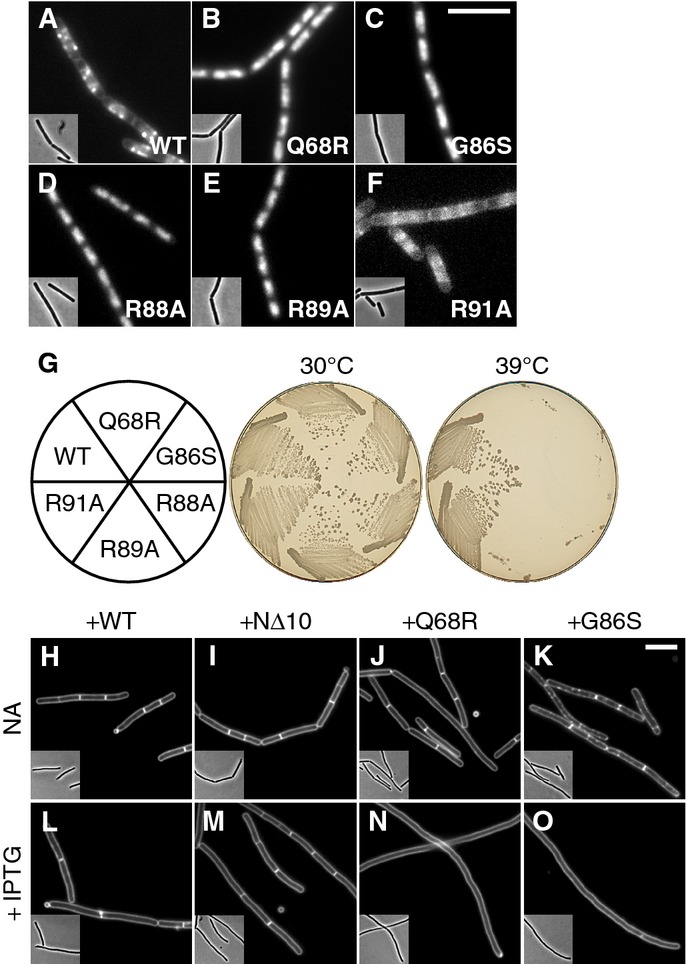
Localisation of ParB-box mutants. Cellular localisation of (A) Noc-mYFP (DWA206), (B) NocQ68R-mYFP (DWA285), (C) NocG86S-mYFP (DWA286), (D) NocR88A-mYFP (DWA545), (E) NocR89A-mYFP (DWA546) and (F) NocR91A-mYFP (DWA547). Insets show the corresponding phase contrast images. Scale bar, 5 μm.
Ability of ParB-box mutants to rescue the growth defect of Δnoc ΔminCD. Strains DWA564 (Pxyl-noc-myfp), 590 (Pxyl-nocQ68R-myfp), 568 (Pxyl-nocG86S-myfp), 598 (Pxyl-nocR88A-myfp), 600 (Pxyl-nocR89A-myfp) and 602 (Pxyl-nocR91A-myfp) were streaked on plates containing 0.5% w/v xylose and incubated for 18 h at either 30 or 39°C, as indicated, before being photographed.
ParB-box mutants are dominant-negative. Cells of strains DWA362 (ΔminCD,Pspac(hy)-noc), 363 (ΔminCD,Pspac(hy)-nocNΔ10), 364 (ΔminCD,Pspac(hy)-nocQ68R) and 365 (ΔminCD,Pspac(hy)-nocG86S) were examined after growth for 2 h at 42°C with either no additions (NA) (H–K) or in the presence of 1 mM IPTG (L–O), as indicated. Cell membranes were stained with FM5-95. Insets show the corresponding phase contrast images. Scale bar, 5 μm.
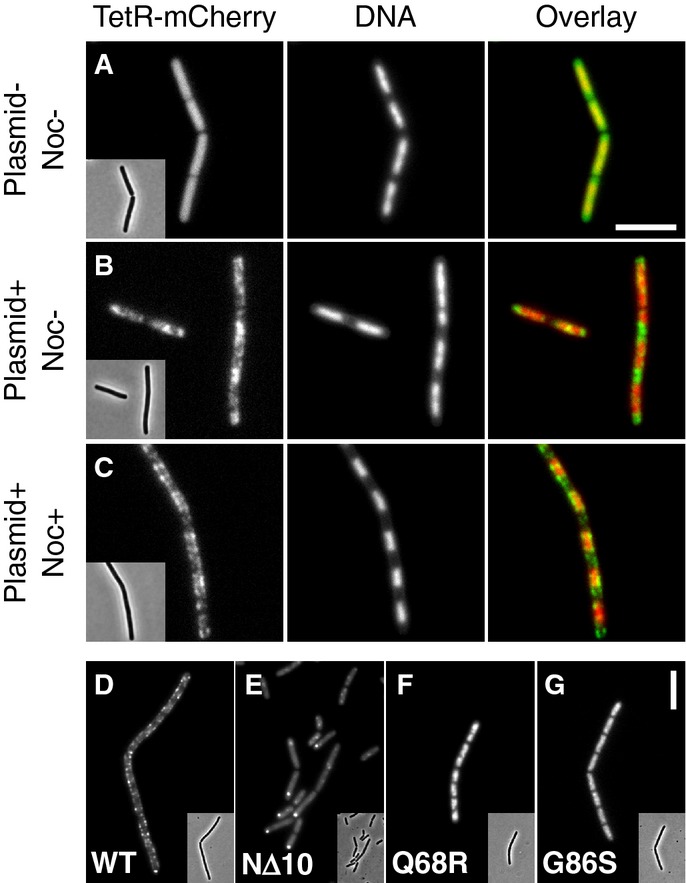
Representative images of TetR-mCherry in cells lacking (A) or containing (B and C) the NBS plasmid pDWA117. Strains DWA427 (A) and 429 (B and C) (both strains contain Pspac-noc) were examined after growth for 2 h in the absence (A and B) and presence (C) of 1 mM IPTG. DNA was stained with DAPI.
Representative images showing the localisation of (D) Noc-mYFP (DWA519), (E) NocNΔ10-mYFP (DWA522), (F) NocQ68R-mYFP (DWA520) and (G) NocG86S-mYFP (DWA521), in the presence of the NBS plasmid pSG4929.
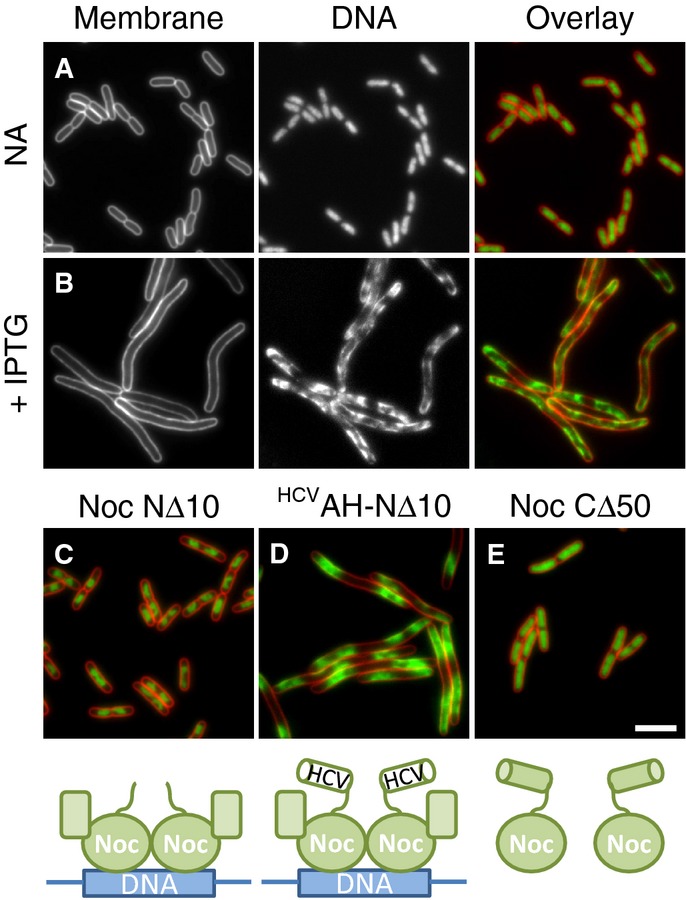
Effects of Noc overproduction on cell division and nucleoid morphology in E. coli. Cells of strain DWA261 carrying pDWA37 (PA1/04/03-noc) were examined after growth in LB with either no additions (A) or after induction for 1 h with 1 mM IPTG (B).
Effects of overproduction of Noc variants on nucleoid morphology. Cells of strains DWA266 (PA1/04/03-nocNΔ10) (C), 270 (PA1/04/03-HCVAH-nocNΔ10) (D) and 267 (PA1/04/03-nocCΔ50) (E) carrying plasmids for the overproduction of the indicated mutants (see cartoons underneath panels) were grown in LB and examined after growth for 1 h in the presence of 1 mM IPTG. Cell membranes and DNA were stained with FM5-95 and DAPI, respectively. Scale bar, 5 μm.
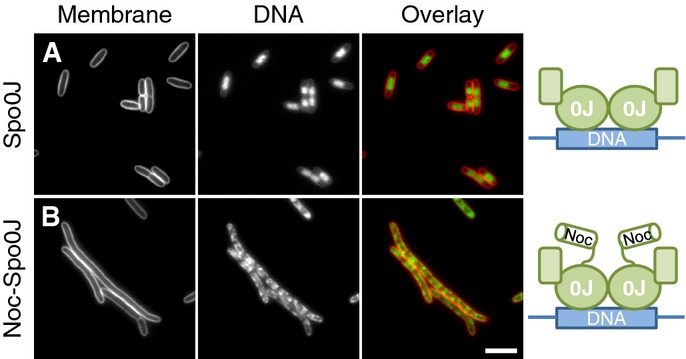
Effects of Spo0J and Noc30-Spo0J overproduction on cell division and nucleoid morphology in E. coli. Cells of strains DWA271 (PA1/04/03-spo0J) (A) and 272 (PA1/04/03-noc-spo0J) (B) were grown in LB in the presence of 1 mM IPTG, to induce the expression of either Spo0J (A) or the Noc-Spo0J hybrid (B), and were examined 1 h post-induction. Cell membranes and DNA were stained with FM5–95 and DAPI, respectively. Scale bar, 5 μm.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
