

Nicotiana small RNA sequences support a host genome origin of cucumber mosaic virus satellite RNA
- PMID: 25568943
- PMCID: PMC4287446
- DOI: 10.1371/journal.pgen.1004906
Nicotiana small RNA sequences support a host genome origin of cucumber mosaic virus satellite RNA
Erratum in
-
Correction: Nicotiana small RNA sequences support a host genome origin of Cucumber mosaic virus satellite RNA.PLoS Genet. 2015 Apr 27;11(4):e1005196. doi: 10.1371/journal.pgen.1005196. eCollection 2015 Apr. PLoS Genet. 2015. PMID: 25915543 Free PMC article.
Abstract
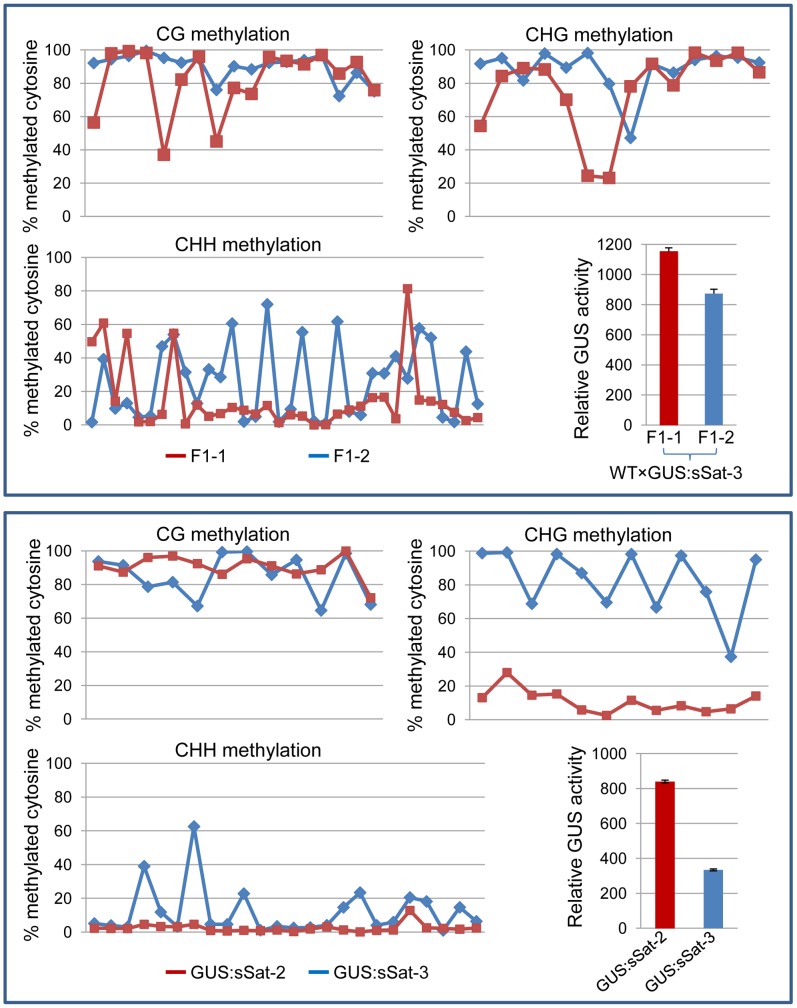
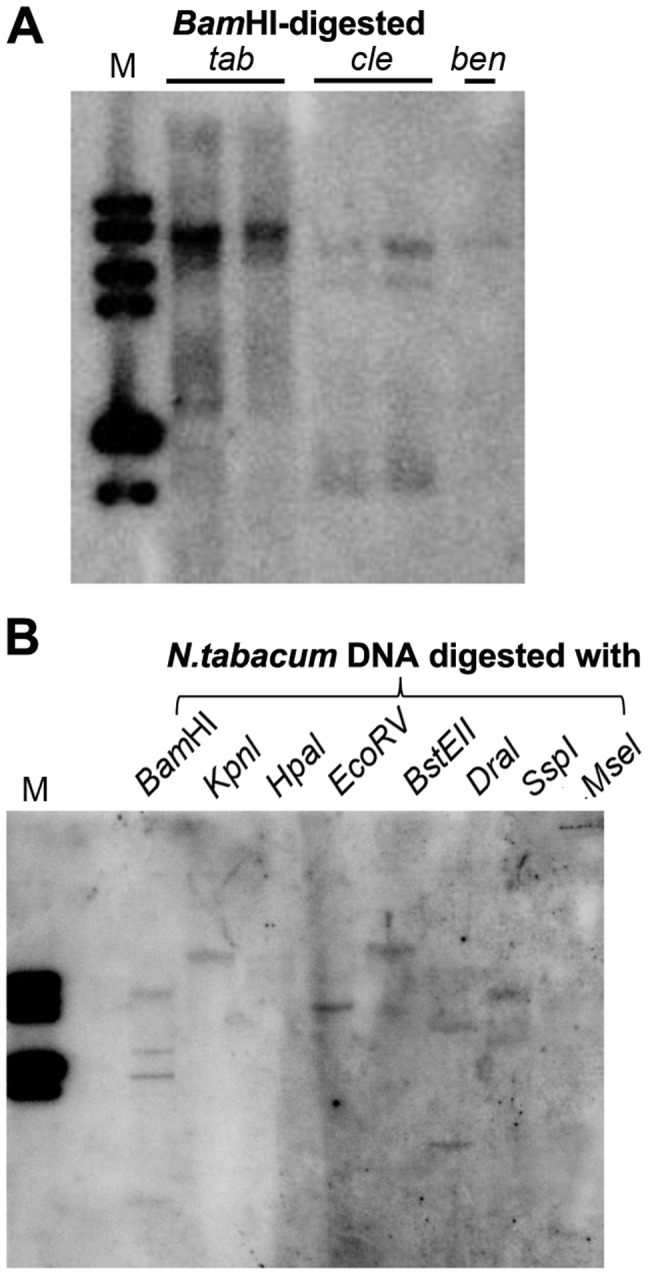
Satellite RNAs (satRNAs) are small noncoding subviral RNA pathogens in plants that depend on helper viruses for replication and spread. Despite many decades of research, the origin of satRNAs remains unknown. In this study we show that a β-glucuronidase (GUS) transgene fused with a Cucumber mosaic virus (CMV) Y satellite RNA (Y-Sat) sequence (35S-GUS:Sat) was transcriptionally repressed in N. tabacum in comparison to a 35S-GUS transgene that did not contain the Y-Sat sequence. This repression was not due to DNA methylation at the 35S promoter, but was associated with specific DNA methylation at the Y-Sat sequence. Both northern blot hybridization and small RNA deep sequencing detected 24-nt siRNAs in wild-type Nicotiana plants with sequence homology to Y-Sat, suggesting that the N. tabacum genome contains Y-Sat-like sequences that give rise to 24-nt sRNAs capable of guiding RNA-directed DNA methylation (RdDM) to the Y-Sat sequence in the 35S-GUS:Sat transgene. Consistent with this, Southern blot hybridization detected multiple DNA bands in Nicotiana plants that had sequence homology to Y-Sat, suggesting that Y-Sat-like sequences exist in the Nicotiana genome as repetitive DNA, a DNA feature associated with 24-nt sRNAs. Our results point to a host genome origin for CMV satRNAs, and suggest novel approach of using small RNA sequences for finding the origin of other satRNAs.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





References
-
- Simon AE, Roossinck MJ, Havelda Z (2004) Plant virus satellite and defective interfering RNAs: new paradigms for a new century. Annu Rev Phytopathol 42: 415–437. - PubMed
-
- Garcia-Arenal F, Palukaitis P (1999) Structure and functional relationships of satellite RNAs of cucumber mosaic virus. Satellites and Defective Viral RNAs: Springer. pp. 37–63. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
