

Preferential use of central metabolism in vivo reveals a nutritional basis for polymicrobial infection
- PMID: 25568946
- PMCID: PMC4287612
- DOI: 10.1371/journal.ppat.1004601
Preferential use of central metabolism in vivo reveals a nutritional basis for polymicrobial infection
Abstract
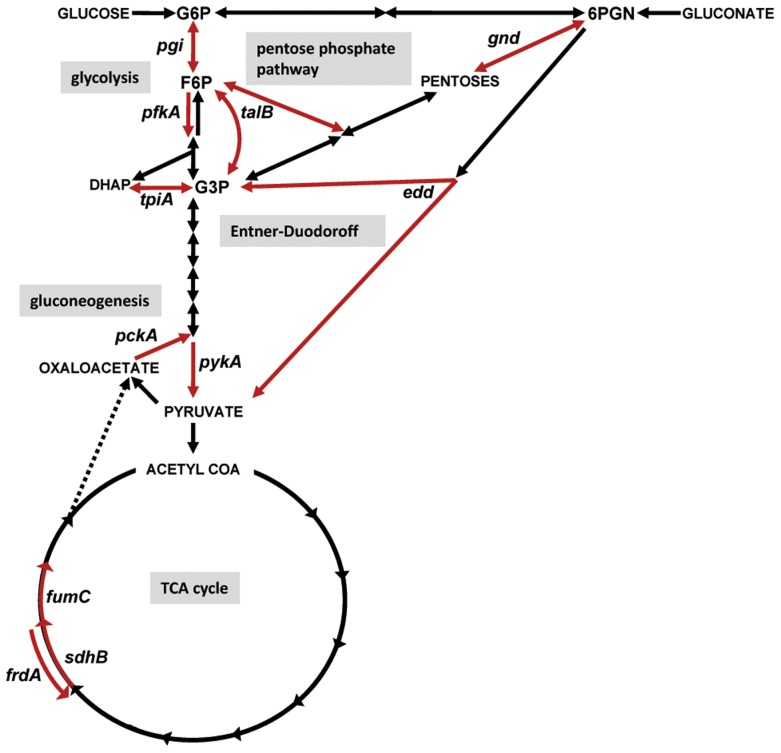
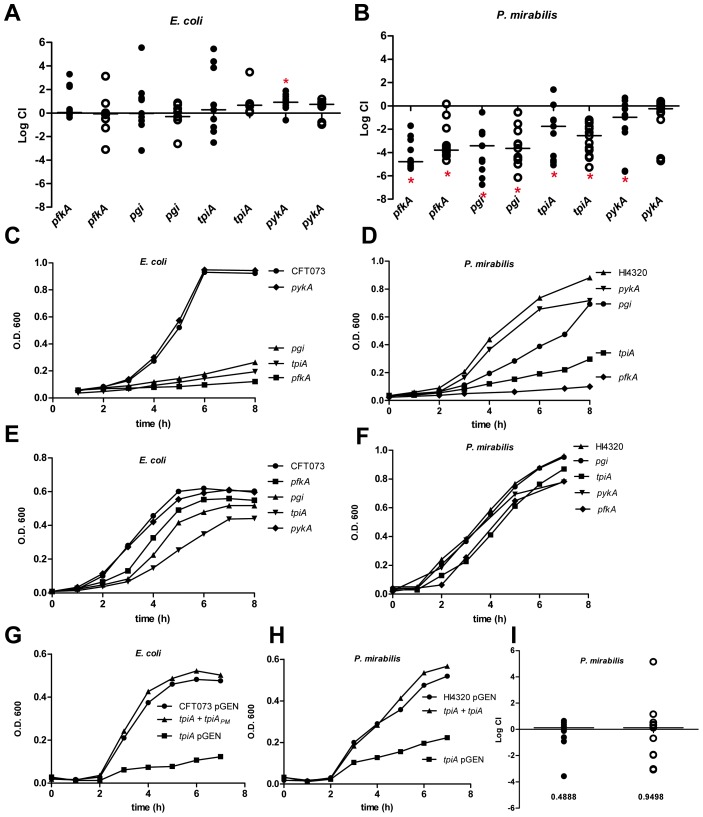
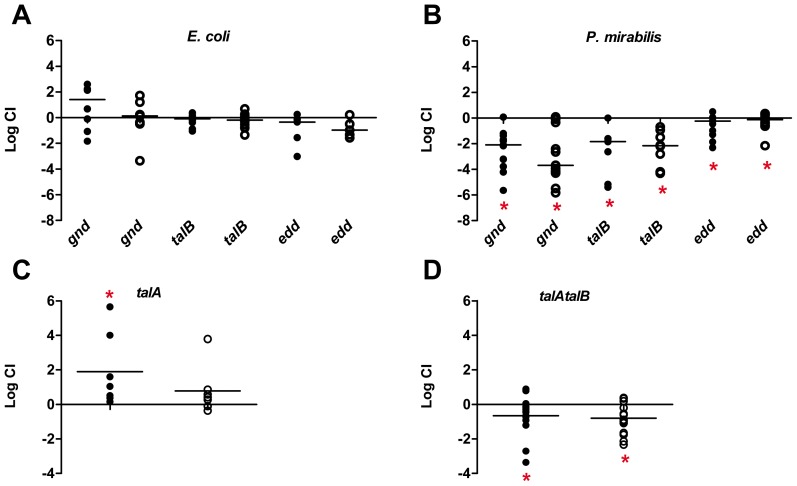
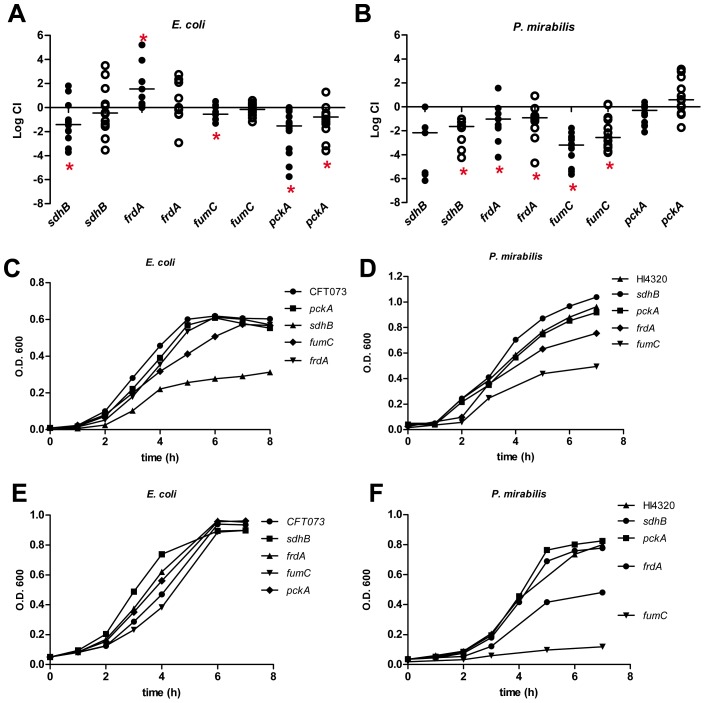
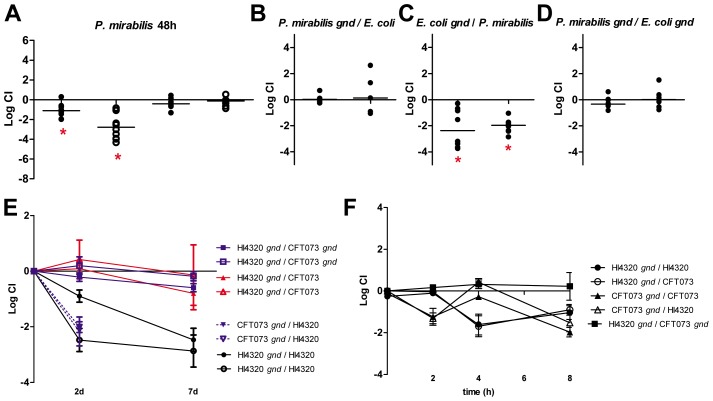
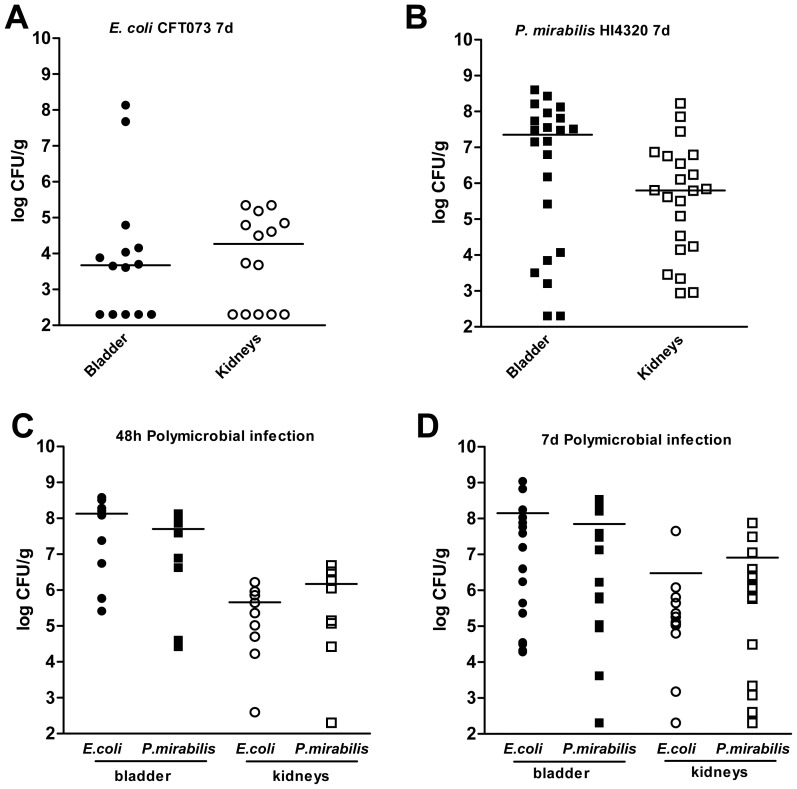
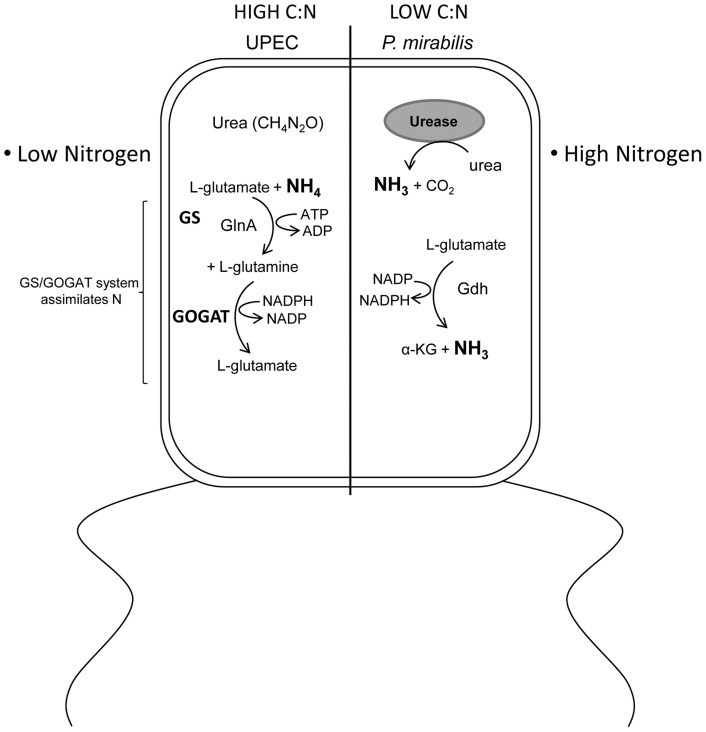
The human genitourinary tract is a common anatomical niche for polymicrobial infection and a leading site for the development of bacteremia and sepsis. Most uncomplicated, community-acquired urinary tract infections (UTI) are caused by Escherichia coli, while another bacterium, Proteus mirabilis, is more often associated with complicated UTI. Here, we report that uropathogenic E. coli and P. mirabilis have divergent requirements for specific central pathways in vivo despite colonizing and occupying the same host environment. Using mutants of specific central metabolism enzymes, we determined glycolysis mutants lacking pgi, tpiA, pfkA, or pykA all have fitness defects in vivo for P. mirabilis but do not affect colonization of E. coli during UTI. Similarly, the oxidative pentose phosphate pathway is required only for P. mirabilis in vivo. In contrast, gluconeogenesis is required only for E. coli fitness in vivo. The remarkable difference in central pathway utilization between E. coli and P. mirabilis during experimental UTI was also observed for TCA cycle mutants in sdhB, fumC, and frdA. The distinct in vivo requirements between these pathogens suggest E. coli and P. mirabilis are not direct competitors within host urinary tract nutritional niche. In support of this, we found that co-infection with E. coli and P. mirabilis wild-type strains enhanced bacterial colonization and persistence of both pathogens during UTI. Our results reveal that complementary utilization of central carbon metabolism facilitates polymicrobial disease and suggests microbial activity in vivo alters the host urinary tract nutritional niche.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Eisenreich W, Dandekar T, Heesemann J, Goebel W (2010) Carbon metabolism of intracellular bacterial pathogens and possible links to virulence. Nat Rev Microbiol 8: 401–412. - PubMed
-
- Poncet S, Milohanic E, Maze A, Nait Abdallah J, Ake F, et al. (2009) Correlations between carbon metabolism and virulence in bacteria. Contrib Microbiol 16: 88–102. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
