

Long non-coding RNA CCAT1 promotes gallbladder cancer development via negative modulation of miRNA-218-5p
- PMID: 25569100
- PMCID: PMC4669740
- DOI: 10.1038/cddis.2014.541
Long non-coding RNA CCAT1 promotes gallbladder cancer development via negative modulation of miRNA-218-5p
Abstract
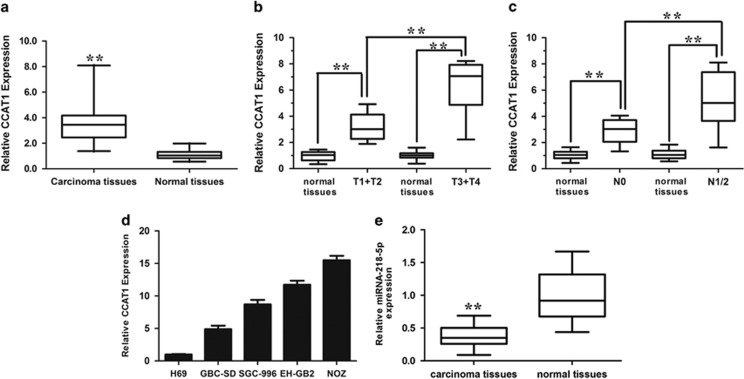
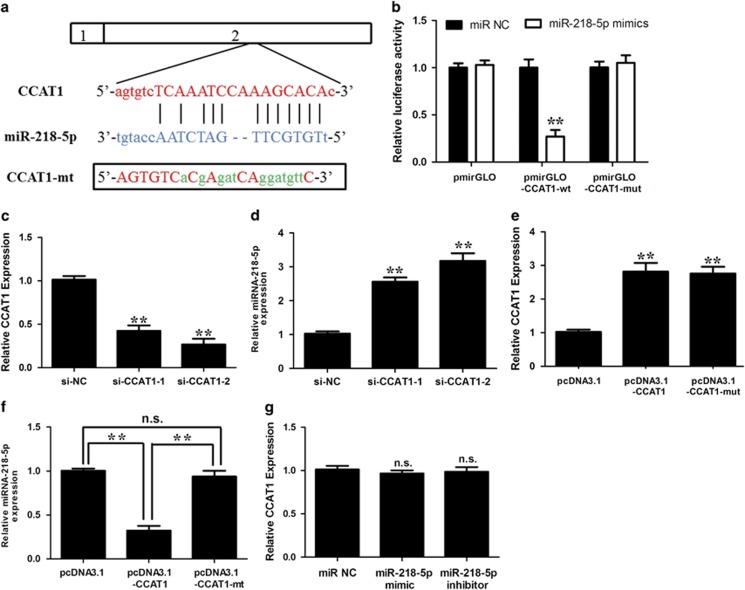
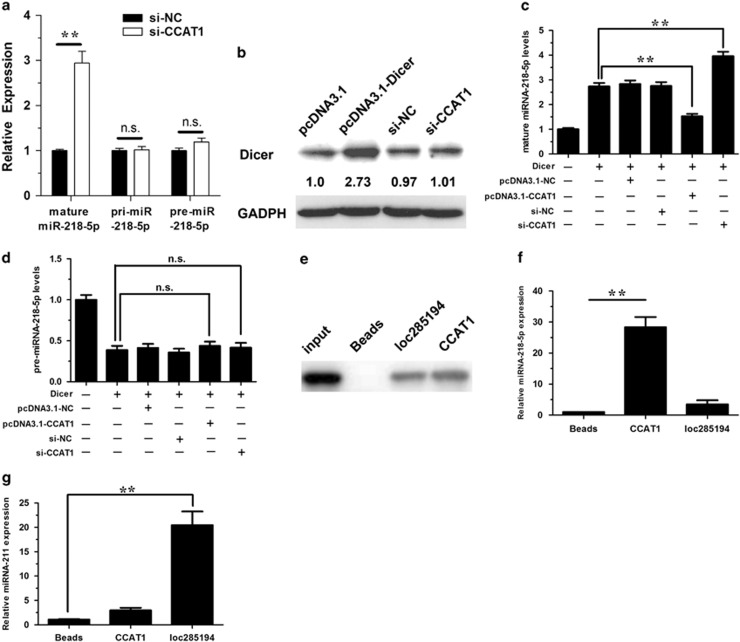
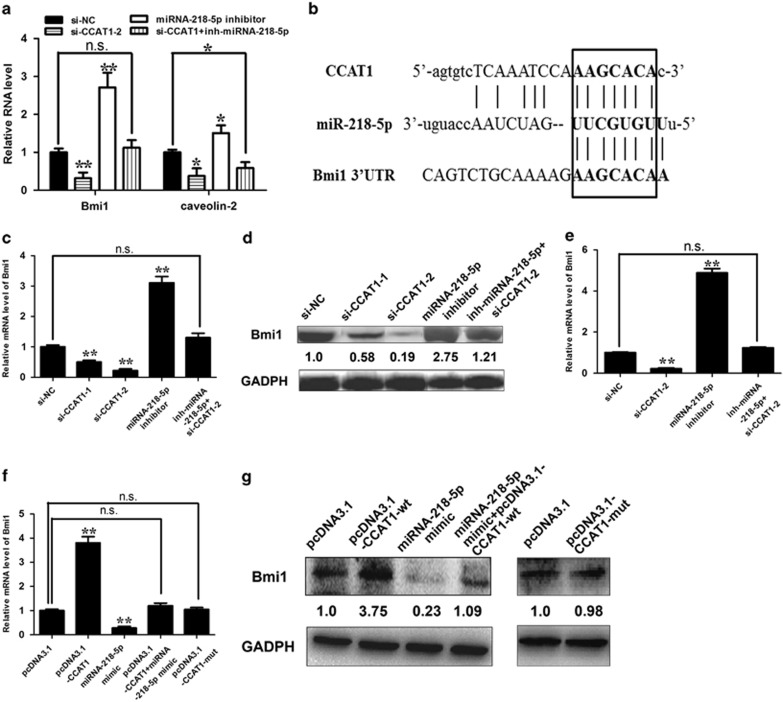
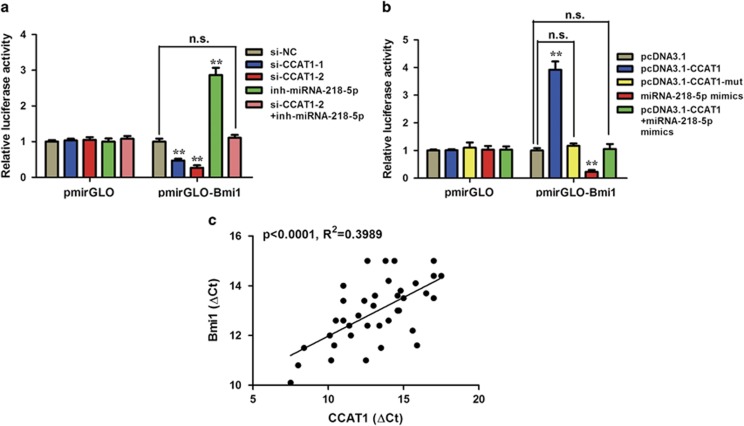
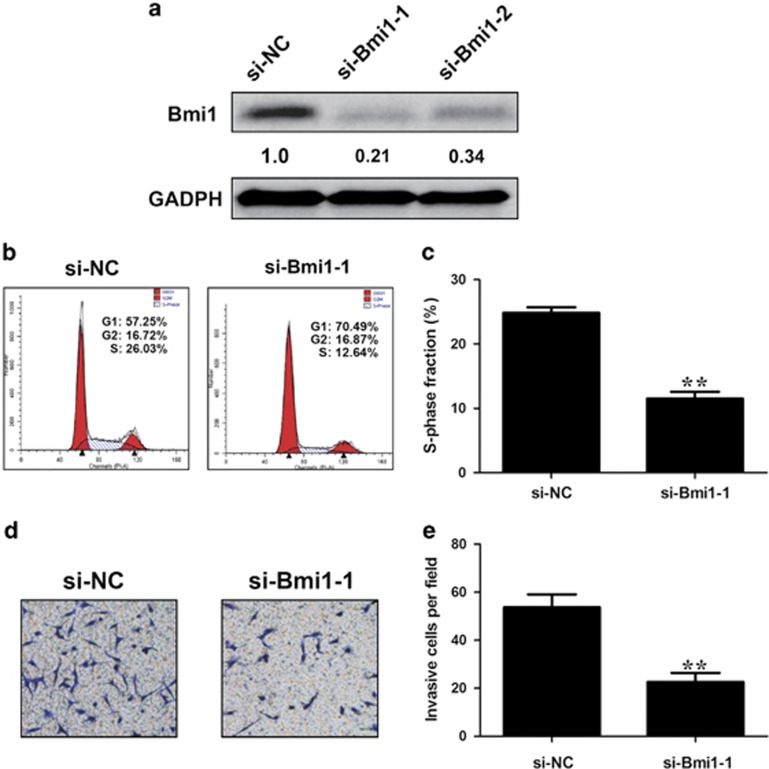
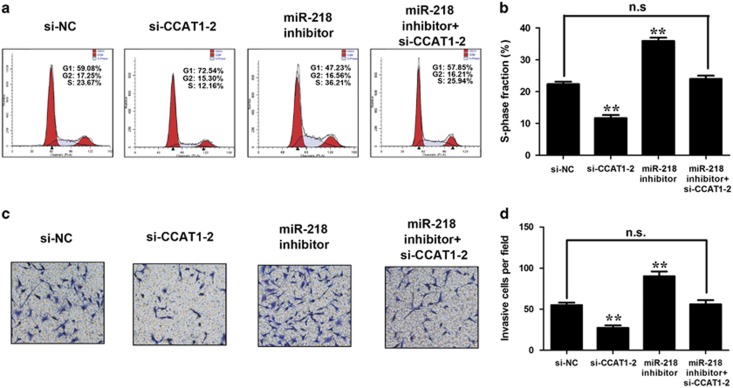
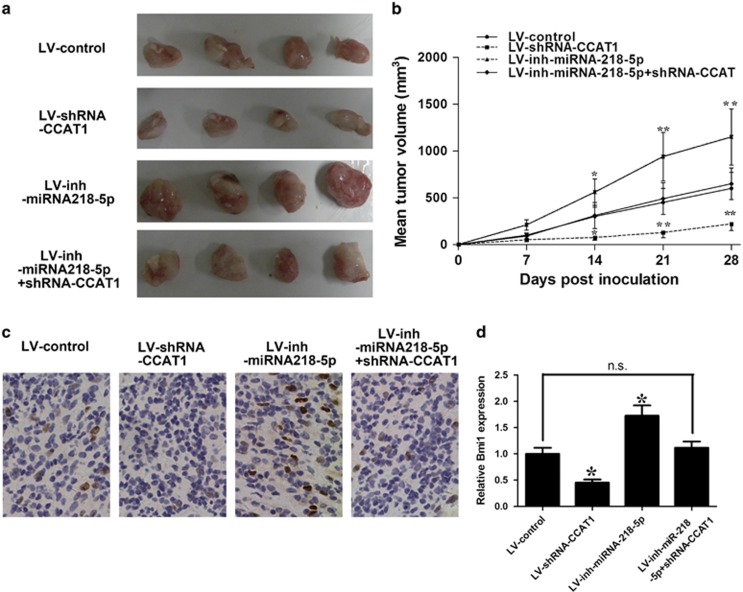
Protein-coding genes account for only ~2% of the human genome, whereas the vast majority of transcripts are non-coding RNAs (ncRNAs) including long ncRNAs (lncRNAs). A growing volume of literature has proposed that lncRNAs are important factors in cancer. Colon cancer-associated transcript-1 (CCAT1), an lncRNA, which was first identified in colon cancer, was previously shown to promote tumor development and be a negative prognostic factor in gastric cancer. However, the mechanism through which CCAT1 exerts its oncogenic activity remains largely unknown. Recently, a novel regulatory mechanism has been proposed in which RNAs can cross-talk with each other via competing shared for microRNAs (miRNAs). The proposed competitive endogenous RNAs could mediate the bioavailability of miRNAs on their targets, thus imposing another level of posttranscriptional regulation. In this study, we demonstrated that CCAT1 was upregulated in gallbladder cancer (GBC) tissues. CCAT1 silencing downregulated, whereas CCAT1 overexpression enhanced the expression of miRNA-218-5p target gene Bmi1 through competitively 'spongeing' miRNA-218-5p. Our data revealed that CCAT1 knockdown impaired the proliferation and invasiveness of GBC cells, at least in part through affecting miRNA-218-5p-mediated regulation of Bmi1. Moreover, CCAT1 transcript level was correlated with Bmi1 mRNA level in GBC tissues. Together, these results suggest that CCAT1 is a driver of malignancy, which acts in part through 'spongeing' miRNA-218-5p.
Figures
References
-
- Ooi A, Suzuki S, Nakazawa K, Itakura J, Imoto I, Nakamura H et al. Gene amplification of Myc and its coamplification with ERBB2 and EGFR in gallbladder adenocarcinoma. Anticancer Res 2009; 29: 19–26. - PubMed
-
- Shi X, Sun M, Liu H, Yao Y, Song Y. Long non-coding RNAs: a new frontier in the study of human diseases. Cancer Lett 2013; 339: 159–166. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
