

Sucrose in cyanobacteria: from a salt-response molecule to play a key role in nitrogen fixation
- PMID: 25569239
- PMCID: PMC4390843
- DOI: 10.3390/life5010102
Sucrose in cyanobacteria: from a salt-response molecule to play a key role in nitrogen fixation
Abstract
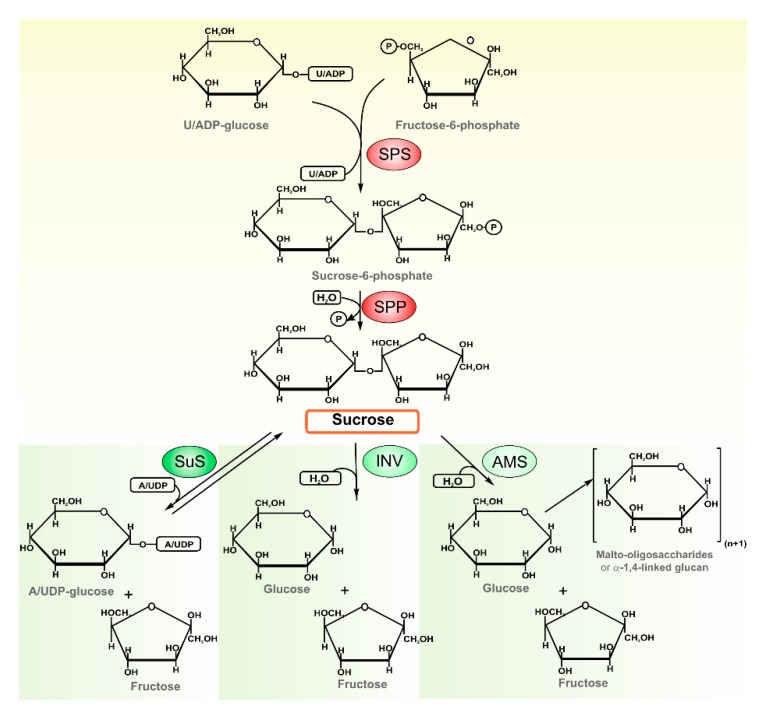
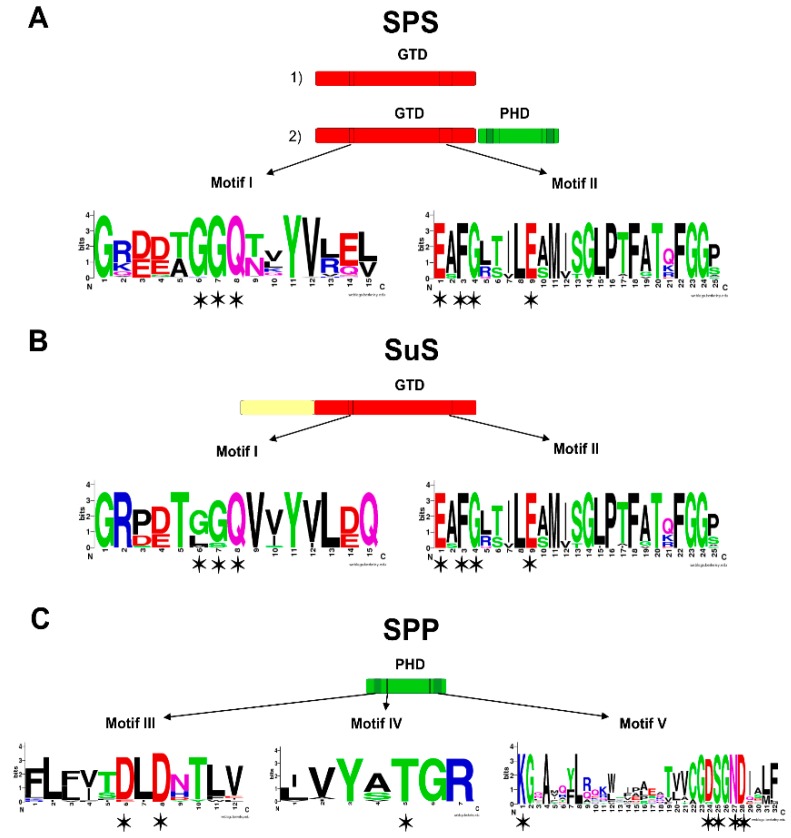
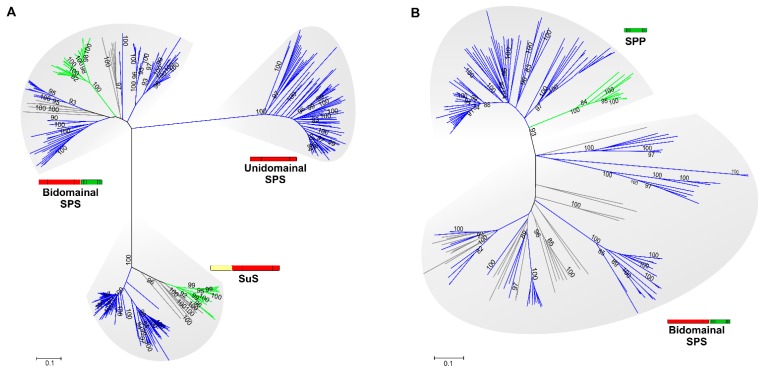
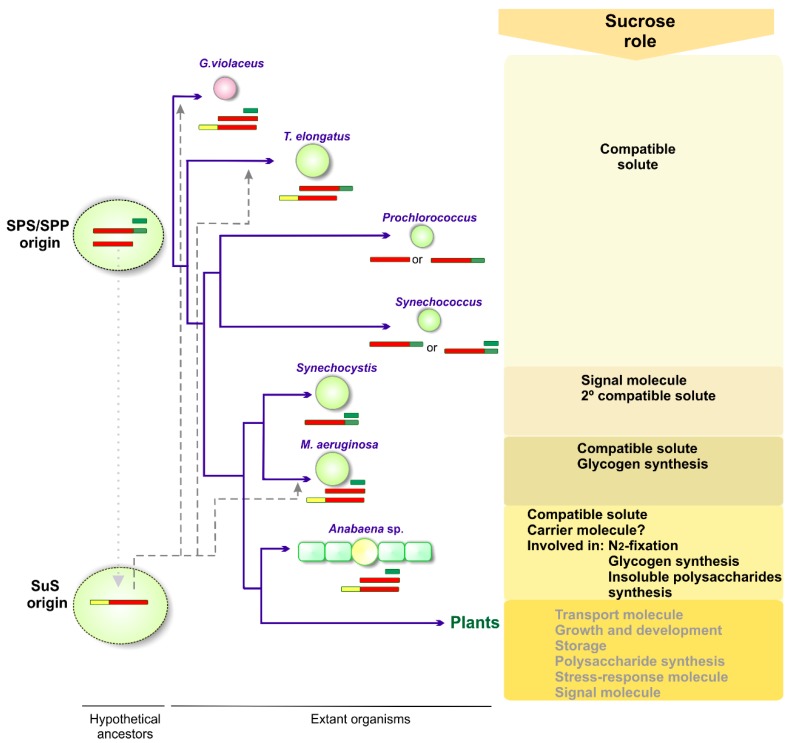
In the biosphere, sucrose is mainly synthesized in oxygenic photosynthetic organisms, such as cyanobacteria, green algae and land plants, as part of the carbon dioxide assimilation pathway. Even though its central position in the functional biology of plants is well documented, much less is known about the role of sucrose in cyanobacteria. In those prokaryotes, sucrose accumulation has been associated with salt acclimation, and considered as a compatible solute in low-salt tolerant strains. In the last years, functional characterizations of sucrose metabolizing enzymes, metabolic control analysis, cellular localization of gene expressions, and reverse genetic experiments have revealed that sucrose metabolism is crucial in the diazotrophic growth of heterocystic strains, and besides, that it can be connected to glycogen synthesis. This article briefly summarizes the current state of knowledge of sucrose physiological functions in modern cyanobacteria and how they might have evolved taking into account the phylogenetic analyses of sucrose enzymes.
Figures
References
-
- Rippka R., Deruelles J., Waterbury J.B., Herdman M., Stanier R.Y. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J. Gen. Microbiol. 1979;111:1–61. doi: 10.1099/00221287-111-1-1. - DOI
-
- Whitton B.A., Potts M. Introduction to the cyanobacteria. In: Whitton B.A., editor. Ecology of Cyanobacteria II. Springer; Dordrecht, The Netherlands: 2012. pp. 1–13.
-
- Reed R.H., Stewart W.D.P. The responses of cyanobacteria to salt stress. In: Rogers L.J., Gallon J.R., editors. Biochemistry of the Algae and Cyanobacteria. Clarendon Press; Oxford, UK: 1988. pp. 217–231. Chapter 12.
-
- Blumwald E., Tel-Or E. Osmoregulation and cell composition in salt-adaptation of Nostoc. Muscorum. Arch. Microbiol. 1982;132:168–172. doi: 10.1007/BF00508725. - DOI
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
