

Heteromers of amyloid precursor protein in cerebrospinal fluid
- PMID: 25573162
- PMCID: PMC4298044
- DOI: 10.1186/1750-1326-10-2
Heteromers of amyloid precursor protein in cerebrospinal fluid
Abstract
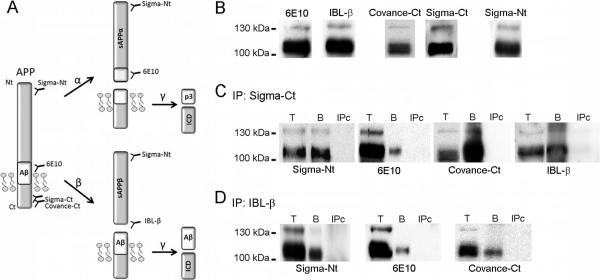
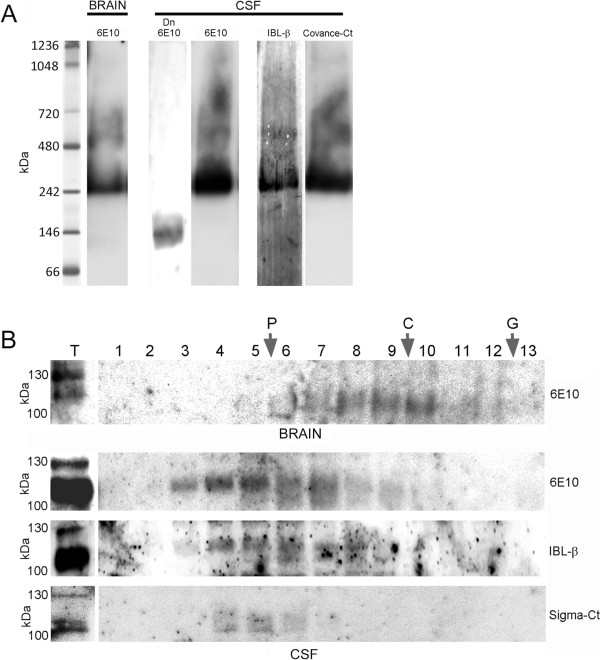
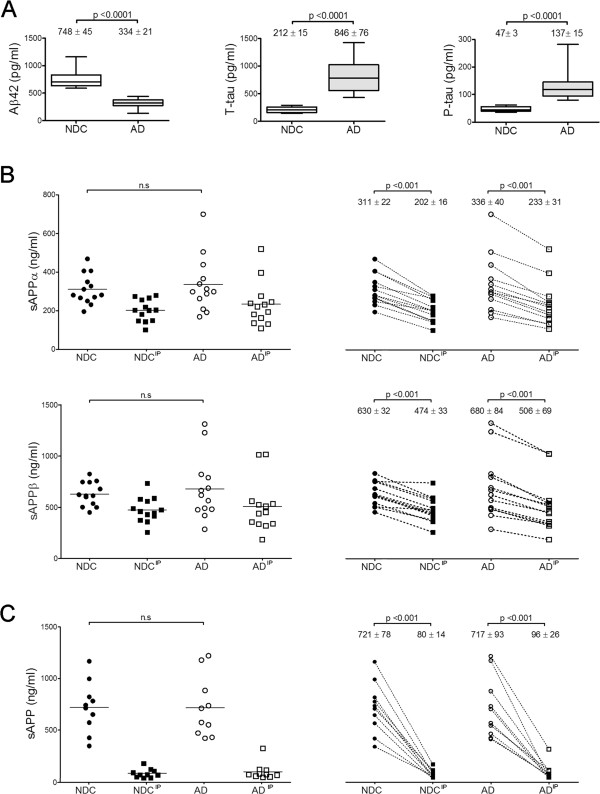
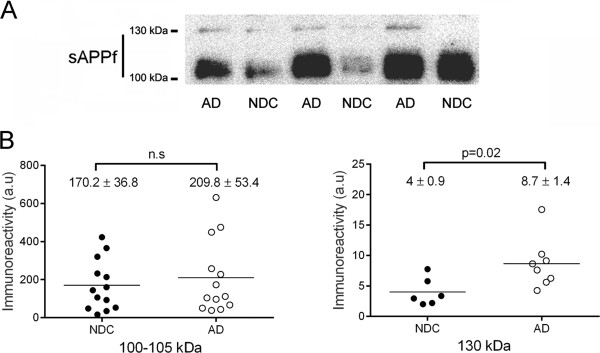
Background: Soluble fragments of the amyloid precursor protein (APP) generated by α- and β-secretases, sAPPα and sAPPβ, have been postulated as promising new cerebrospinal fluid (CSF) biomarkers for the clinical diagnosis of Alzheimer's disease (AD). However, the capacity of these soluble proteins to assemble has not been explored and could be relevant. Our aim is to characterize possible sAPP oligomers that could contribute to the quantification of sAPPα and sAPPβ in CSF by ELISA, as well as to characterize the possible presence of soluble full-length APP (sAPPf).
Results: We employed co-immunoprecipitation, native polyacrylamide gel electrophoresis and ultracentrifugation in sucrose density gradients to characterize sAPP oligomers in CSF. We have characterized the presence of sAPPf in CSF from NDC and AD subjects and demonstrated that all forms, including sAPPα and sAPPβ, are capable of assembling into heteromers, which differ from brain APP membrane-dimers. We measured sAPPf, sAPPα and sAPPβ by ELISA in CSF samples from AD (n = 13) and non-disease subjects (NDC, n = 13) before and after immunoprecipitation with antibodies against the C-terminal APP or against sAPPβ. We demonstrated that these sAPP heteromers participate in the quantification of sAPPα and sAPPβ by ELISA. Immunoprecipitation with a C-terminal antibody to remove sAPPf reduced by ~30% the determinations of sAPPα and sAPPβ by ELISA, whereas immunoprecipitation with an APPβ antibody reduced by ~80% the determination of sAPPf and sAPPα.
Conclusions: The presence of sAPPf and sAPP heteromers should be taken into consideration when exploring the levels of sAPPα and sAPPβ as potential CSF biomarkers.
Figures
Similar articles
-
Amyloid precursor protein glycosylation is altered in the brain of patients with Alzheimer's disease.Alzheimers Res Ther. 2020 Aug 12;12(1):96. doi: 10.1186/s13195-020-00664-9. Alzheimers Res Ther. 2020. PMID: 32787955 Free PMC article.
-
Soluble amyloid precursor protein α and β in CSF in Alzheimer's disease.Brain Res. 2013 Jun 4;1513:117-26. doi: 10.1016/j.brainres.2013.03.019. Epub 2013 Mar 27. Brain Res. 2013. PMID: 23541617
-
CNS amyloid-β, soluble APP-α and -β kinetics during BACE inhibition.J Neurosci. 2014 Jun 11;34(24):8336-46. doi: 10.1523/JNEUROSCI.0540-14.2014. J Neurosci. 2014. PMID: 24920637 Free PMC article.
-
Beta-site amyloid precursor protein cleaving enzyme 1 (BACE1) as a biological candidate marker of Alzheimer's disease.Scand J Clin Lab Invest. 2009;69(1):8-12. doi: 10.1080/00365510701864610. Scand J Clin Lab Invest. 2009. PMID: 18609117 Review.
-
Functions of Aβ, sAPPα and sAPPβ : similarities and differences.J Neurochem. 2012 Jan;120 Suppl 1:99-108. doi: 10.1111/j.1471-4159.2011.07584.x. Epub 2011 Dec 7. J Neurochem. 2012. PMID: 22150401 Review.
Cited by
-
Levels of ADAM10 are reduced in Alzheimer's disease CSF.J Neuroinflammation. 2018 Jul 25;15(1):213. doi: 10.1186/s12974-018-1255-9. J Neuroinflammation. 2018. PMID: 30045733 Free PMC article.
-
CSF sAPPα and sAPPβ levels in Alzheimer's Disease and Multiple Other Neurodegenerative Diseases: A Network Meta-Analysis.Neuromolecular Med. 2020 Mar;22(1):45-55. doi: 10.1007/s12017-019-08561-7. Epub 2019 Aug 14. Neuromolecular Med. 2020. PMID: 31414383
-
Soluble APP-α and APP-β in cerebrospinal fluid as potential biomarkers for differential diagnosis of mild cognitive impairment.Aging Clin Exp Res. 2022 Feb;34(2):341-347. doi: 10.1007/s40520-021-01935-7. Epub 2021 Jul 20. Aging Clin Exp Res. 2022. PMID: 34283410
-
Plasma biomarkers of the amyloid pathway are associated with geographic atrophy secondary to age-related macular degeneration.PLoS One. 2020 Aug 7;15(8):e0236283. doi: 10.1371/journal.pone.0236283. eCollection 2020. PLoS One. 2020. PMID: 32764794 Free PMC article.
-
Amyloid precursor protein glycosylation is altered in the brain of patients with Alzheimer's disease.Alzheimers Res Ther. 2020 Aug 12;12(1):96. doi: 10.1186/s13195-020-00664-9. Alzheimers Res Ther. 2020. PMID: 32787955 Free PMC article.
References
-
- Palmert MR, Siedlak SL, Podlisny MB, Greenberg B, Shelton ER, Chan HW, Usiak M, Selkoe DJ, Perry G, Younkin SG. The beta-amyloid protein precursor of Alzheimer disease has soluble derivatives found in human brain and cerebrospinal fluid. Proc. Natl. Acad. Sci. U. S. A. 1989;86:6338–6342. doi: 10.1073/pnas.86.16.6338. - DOI - PMC - PubMed
-
- Ghiso J, Tagliavini F, Timmers WF, Frangione B. Alzheimer’s disease amyloid precursor protein is present in senile plaques and cerebrospinal fluid: immunohistochemical and biochemical characterization. Biochem. Biophys. Res. Commun. 1989;163:430–437. doi: 10.1016/0006-291X(89)92154-2. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
