

Enzyme dynamics from NMR spectroscopy
- PMID: 25574774
- PMCID: PMC4334254
- DOI: 10.1021/ar500340a
Enzyme dynamics from NMR spectroscopy
Abstract
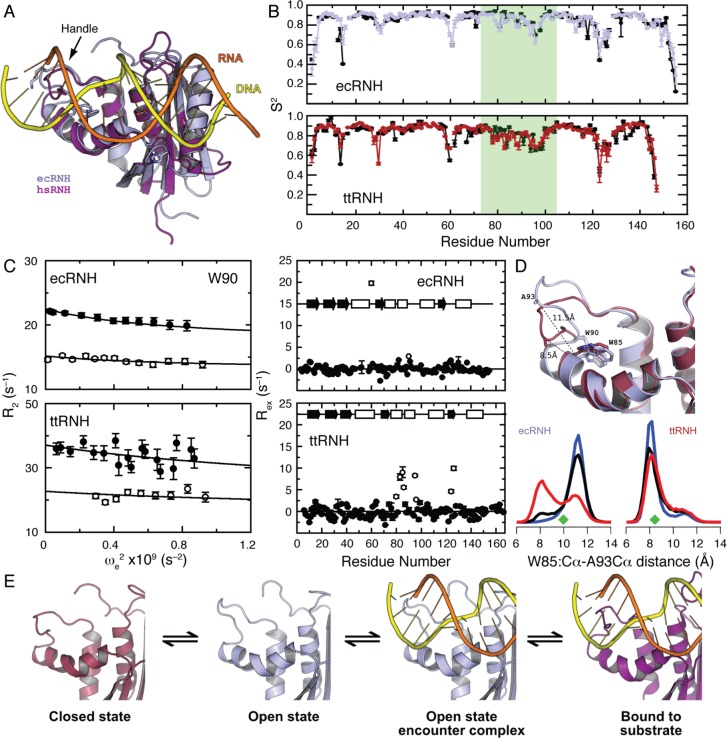
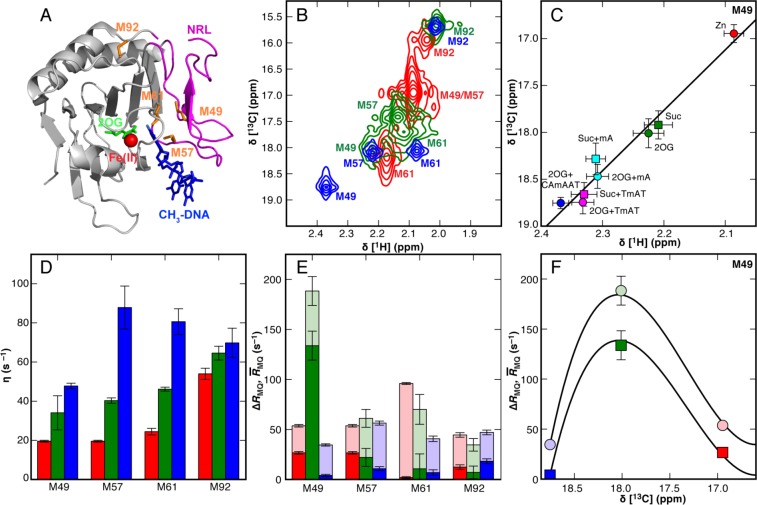
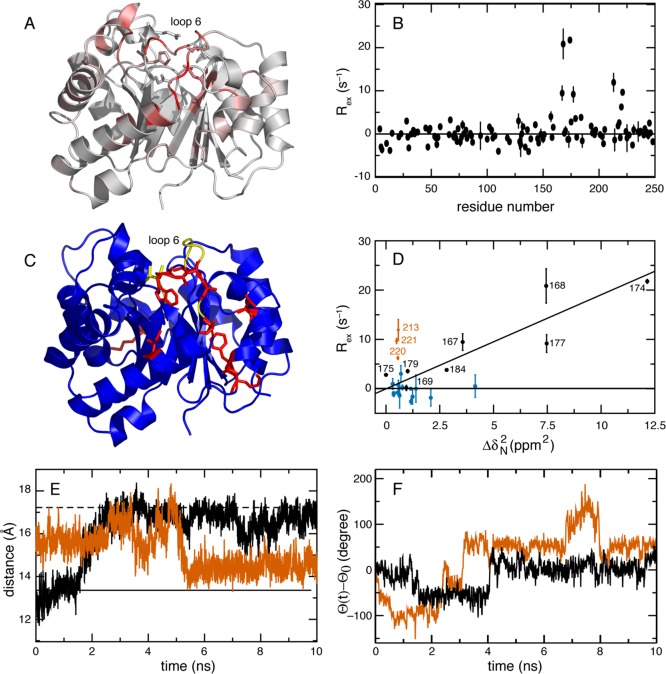
CONSPECTUS: Biological activities of enzymes, including regulation or coordination of mechanistic stages preceding or following the chemical step, may depend upon kinetic or equilibrium changes in protein conformations. Exchange of more open or flexible conformational states with more closed or constrained states can influence inhibition, allosteric regulation, substrate recognition, formation of the Michaelis complex, side reactions, and product release. NMR spectroscopy has long been applied to the study of conformational dynamic processes in enzymes because these phenomena can be characterized over multiple time scales with atomic site resolution. Laboratory-frame spin-relaxation measurements, sensitive to reorientational motions on picosecond-nanosecond time scales, and rotating-frame relaxation-dispersion measurements, sensitive to chemical exchange processes on microsecond-millisecond time scales, provide information on both conformational distributions and kinetics. This Account reviews NMR spin relaxation studies of the enzymes ribonuclease HI from mesophilic (Escherichia coli) and thermophilic (Thermus thermophilus) bacteria, E. coli AlkB, and Saccharomyces cerevisiae triosephosphate isomerase to illustrate the contributions of conformational flexibility and dynamics to diverse steps in enzyme mechanism. Spin relaxation measurements and molecular dynamics (MD) simulations of the bacterial ribonuclease H enzymes show that the handle region, one of three loop regions that interact with substrates, interconverts between two conformations. Comparison of these conformations with the structure of the complex between Homo sapiens ribonuclease H and a DNA:RNA substrate suggests that the more closed state is inhibitory to binding. The large population of the closed conformation in T. thermophilus ribonuclease H contributes to the increased Michaelis constant compared with the E. coli enzyme. NMR spin relaxation and fluorescence spectroscopy have characterized a conformational transition in AlkB between an open state, in which the side chains of methionine residues in the active site are disordered, and a closed state, in which these residues are ordered. The open state is highly populated in the AlkB/Zn(II) complex, and the closed state is highly populated in the AlkB/Zn(II)/2OG/substrate complex, in which 2OG is the 2-oxoglutarate cosubstrate and the substrate is a methylated DNA oligonucleotide. The equilibrium is shifted to approximately equal populations of the two conformations in the AlkB/Zn(II)/2OG complex. The conformational shift induced by 2OG ensures that 2OG binds to AlkB/Zn(II) prior to the substrate. In addition, the opening rate of the closed conformation limits premature release of substrate, preventing generation of toxic side products by reaction with water. Closure of active site loop 6 in triosephosphate isomerase is critical for forming the Michaelis complex, but reopening of the loop after the reaction is (partially) rate limiting. NMR spin relaxation and MD simulations of triosephosphate isomerase in complex with glycerol 3-phosphate demonstrate that closure of loop 6 is a highly correlated rigid-body motion. The MD simulations also indicate that motions of Gly173 in the most flexible region of loop 6 contribute to opening of the active site loop for product release. Considered together, these three enzyme systems illustrate the power of NMR spin relaxation investigations in providing global insights into the role of conformational dynamic processes in the mechanisms of enzymes from initial activation to final product release.
Figures
References
-
- Palmer A. G.NMR spectroscopy: NMR relaxation methods. In Biophysical Techniques for Structural Characterization of Macromolecules; Dyson H. J., Ed.; Academic Press: Oxford, 2012; Vol. 1; pp 216–244.
-
- Palmer A. G.; Massi F. Characterization of the dynamics of biomacromolecules using rotating-frame spin relaxation NMR spectroscopy. Chem. Rev. 2006, 106, 1700–1719. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
