

Independent evolution of four heme peroxidase superfamilies
- PMID: 25575902
- PMCID: PMC4420034
- DOI: 10.1016/j.abb.2014.12.025
Independent evolution of four heme peroxidase superfamilies
Abstract
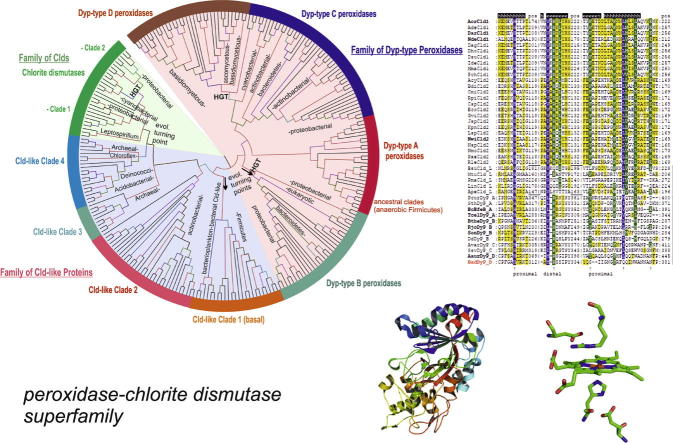
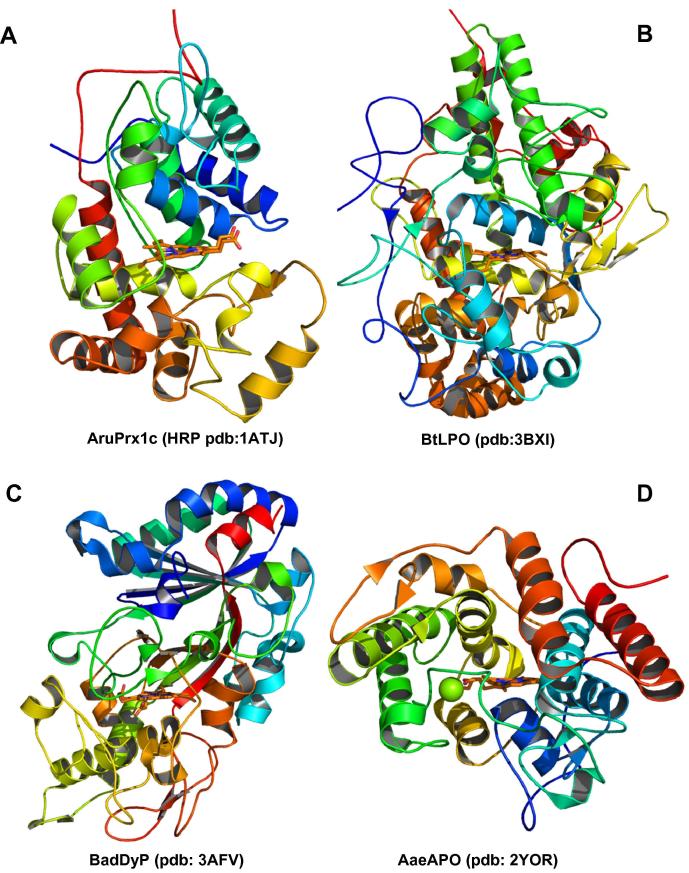
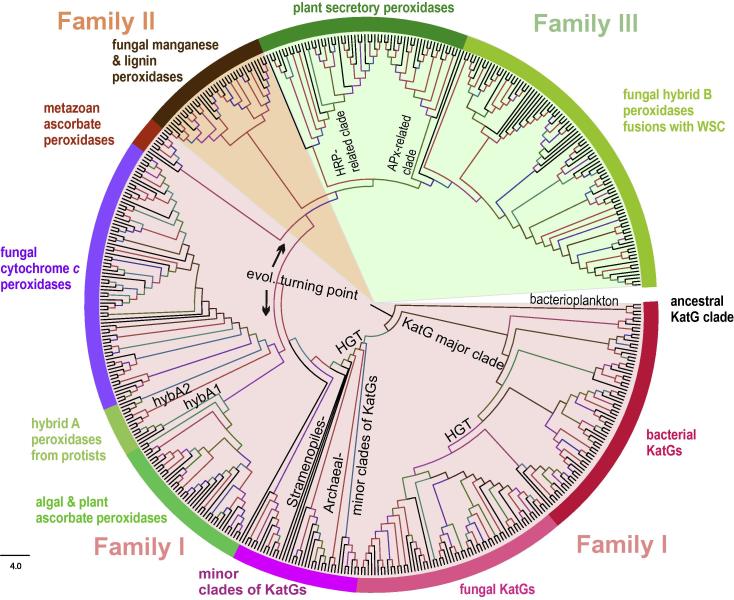
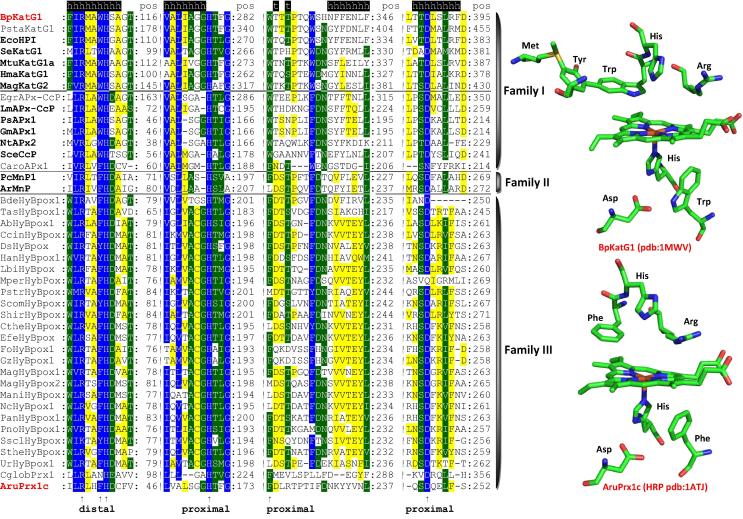
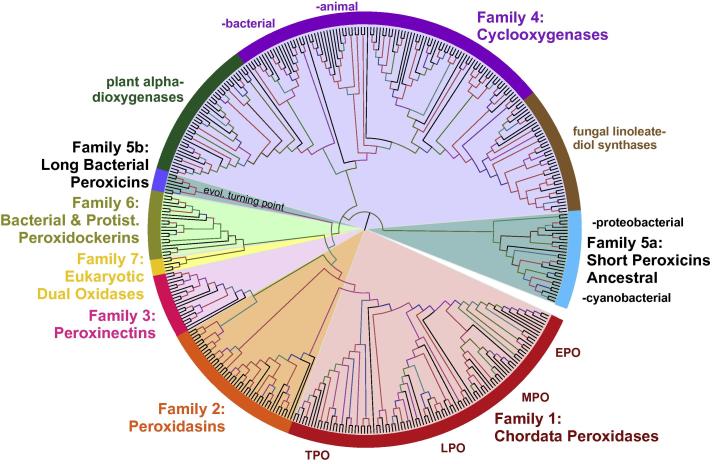
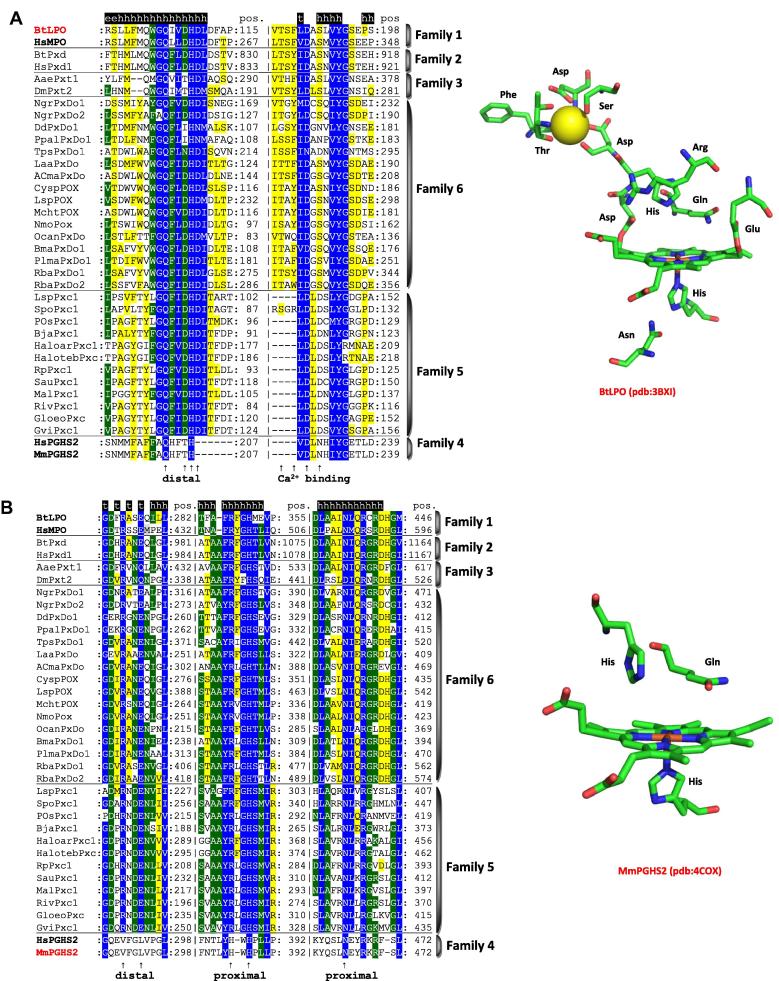
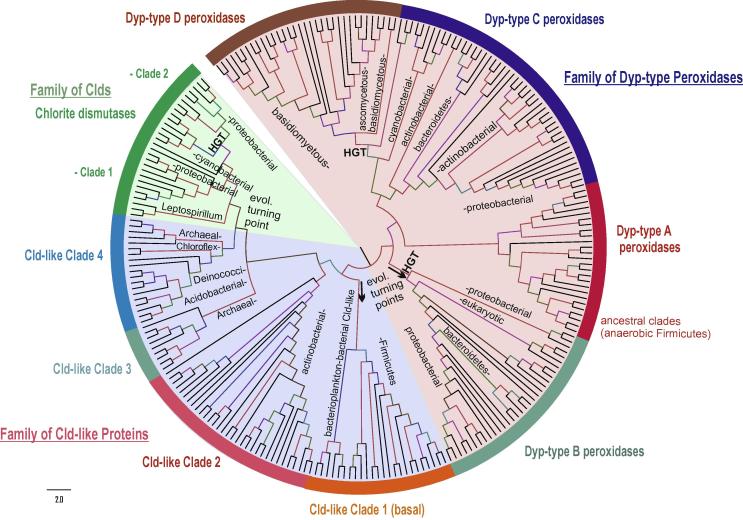
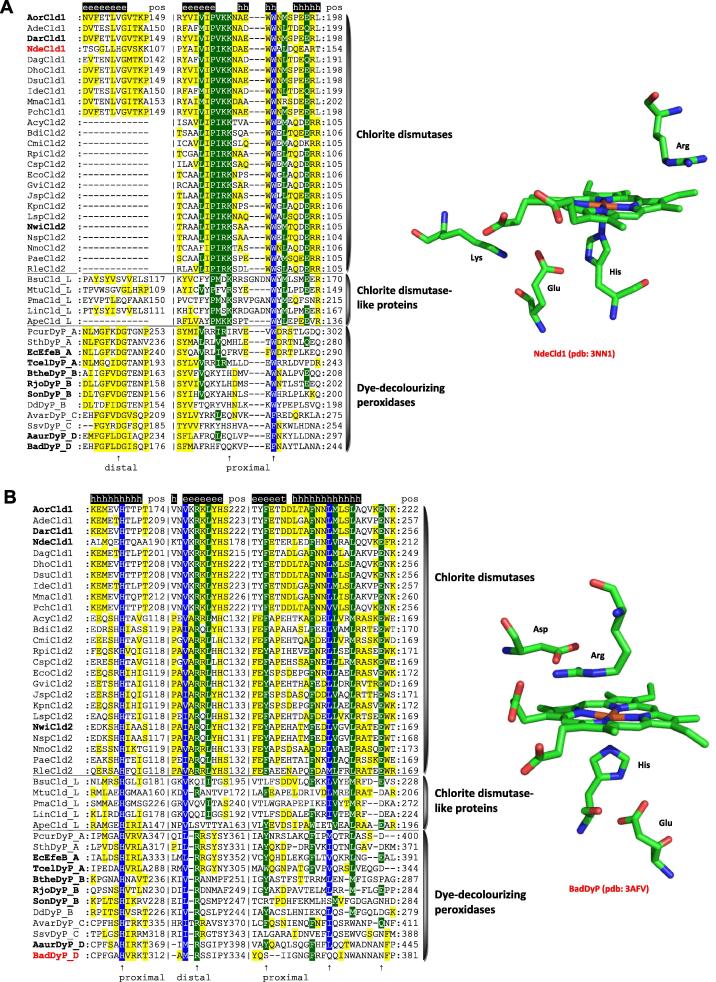
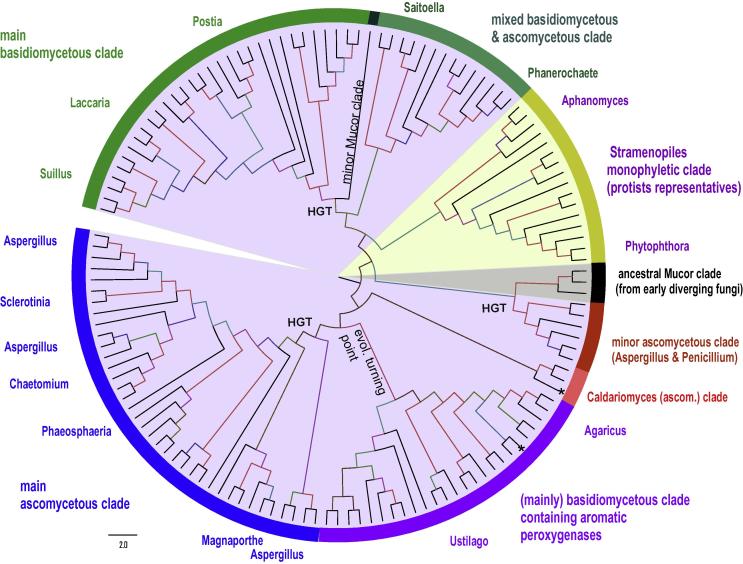
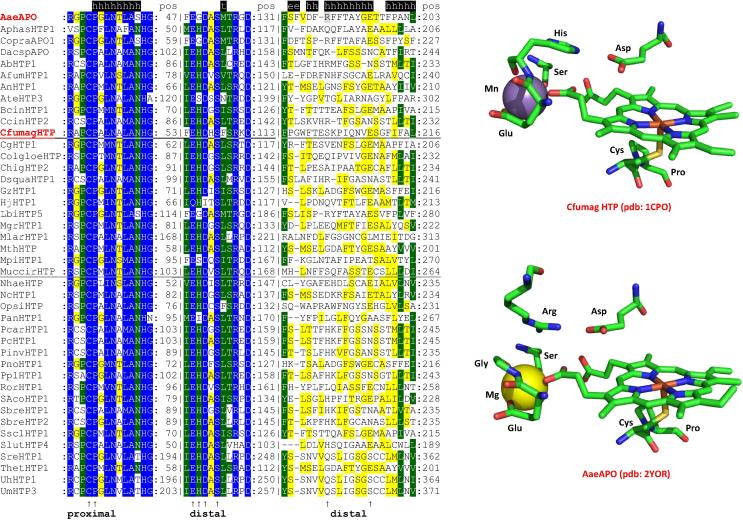
Four heme peroxidase superfamilies (peroxidase-catalase, peroxidase-cyclooxygenase, peroxidase-chlorite dismutase and peroxidase-peroxygenase superfamily) arose independently during evolution, which differ in overall fold, active site architecture and enzymatic activities. The redox cofactor is heme b or posttranslationally modified heme that is ligated by either histidine or cysteine. Heme peroxidases are found in all kingdoms of life and typically catalyze the one- and two-electron oxidation of a myriad of organic and inorganic substrates. In addition to this peroxidatic activity distinct (sub)families show pronounced catalase, cyclooxygenase, chlorite dismutase or peroxygenase activities. Here we describe the phylogeny of these four superfamilies and present the most important sequence signatures and active site architectures. The classification of families is described as well as important turning points in evolution. We show that at least three heme peroxidase superfamilies have ancient prokaryotic roots with several alternative ways of divergent evolution. In later evolutionary steps, they almost always produced highly evolved and specialized clades of peroxidases in eukaryotic kingdoms with a significant portion of such genes involved in coding various fusion proteins with novel physiological functions.
Keywords: Heme peroxidase; Peroxidase–catalase superfamily; Peroxidase–chlorite dismutase superfamily; Peroxidase–cyclooxygenase superfamily; Peroxidase–peroxygenase superfamily.
Copyright © 2015 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Sammut S.J., Finn R.D., Bateman A. Brief. Bioinf. 2008;9:210–219. - PubMed
-
- Chothia C., Cough J. Biochem. J. 2009;419:15–28. - PubMed
-
- Caetano-Anolles G., Wang M., Caetano-Anolles D., Mittenthal J.E. Biochem. J. 2009;417:621–637. - PubMed
-
- Zamocky M., Jakopitsch C., Furtmüller P.G., Dunand C., Obinger C. Proteins. 2008;72:589–605. - PubMed
-
- Welinder K.G. Curr. Opin. Struct. Biol. 1992;2:388–393.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
