

Regulation of splicing factors by alternative splicing and NMD is conserved between kingdoms yet evolutionarily flexible
- PMID: 25576366
- PMCID: PMC4379411
- DOI: 10.1093/molbev/msv002
Regulation of splicing factors by alternative splicing and NMD is conserved between kingdoms yet evolutionarily flexible
Abstract
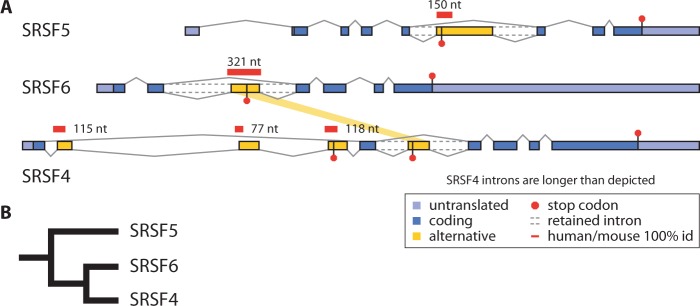
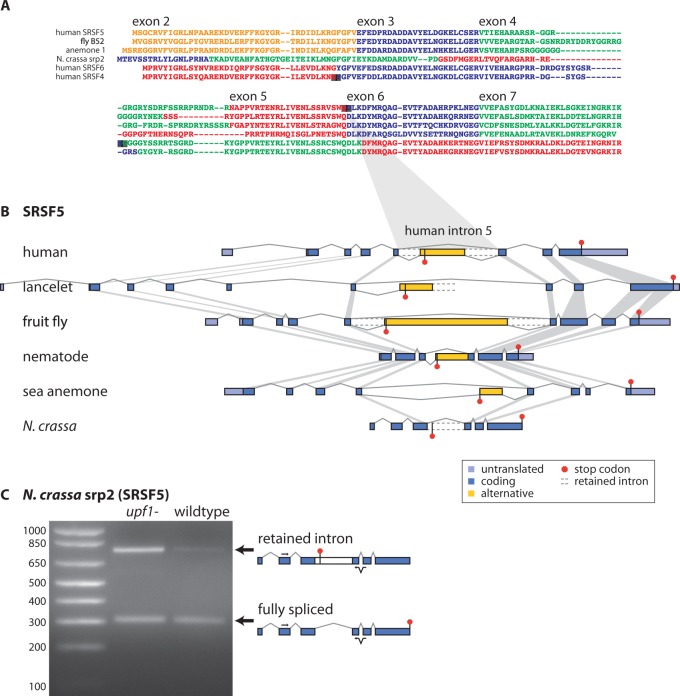
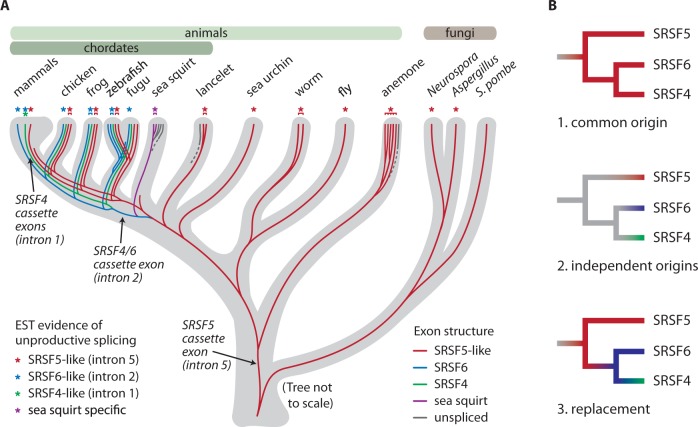
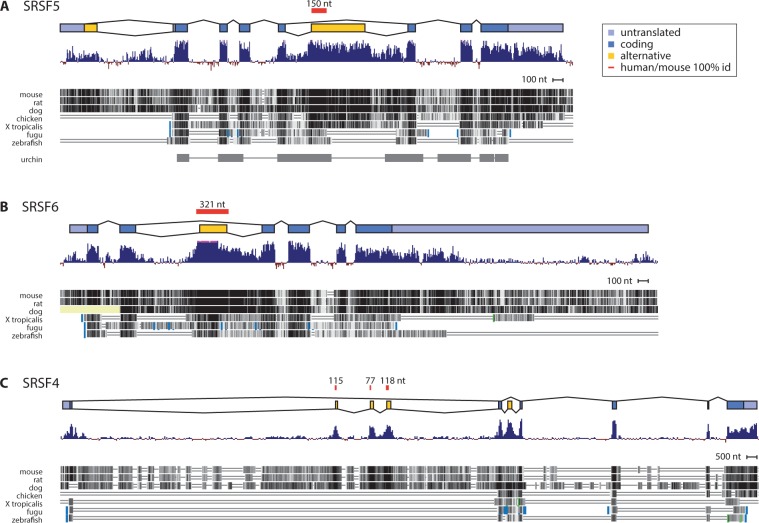
Ultraconserved elements, unusually long regions of perfect sequence identity, are found in genes encoding numerous RNA-binding proteins including arginine-serine rich (SR) splicing factors. Expression of these genes is regulated via alternative splicing of the ultraconserved regions to yield mRNAs that are degraded by nonsense-mediated mRNA decay (NMD), a process termed unproductive splicing (Lareau et al. 2007; Ni et al. 2007). As all human SR genes are affected by alternative splicing and NMD, one might expect this regulation to have originated in an early SR gene and persisted as duplications expanded the SR family. But in fact, unproductive splicing of most human SR genes arose independently (Lareau et al. 2007). This paradox led us to investigate the origin and proliferation of unproductive splicing in SR genes. We demonstrate that unproductive splicing of the splicing factor SRSF5 (SRp40) is conserved among all animals and even observed in fungi; this is a rare example of alternative splicing conserved between kingdoms, yet its effect is to trigger mRNA degradation. As the gene duplicated, the ancient unproductive splicing was lost in paralogs, and distinct unproductive splicing evolved rapidly and repeatedly to take its place. SR genes have consistently employed unproductive splicing, and while it is exceptionally conserved in some of these genes, turnover in specific events among paralogs shows flexible means to the same regulatory end.
Keywords: alternative splicing; nonsense mediated decay; ultraconserved elements.
© The Author 2015. Published by Oxford University Press on behalf of the society for Molecular Biology and Evolution.
Figures
References
-
- Änkö ML, Morales L, Henry I, Beyer A, Neugebauer KM. Global analysis reveals SRp20- and SRp75-specific mRNPs in cycling and neural cells. Nat Struct Mol Biol. 2010;17:962–970. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
