

DNA double-strand breaks and telomeres play important roles in trypanosoma brucei antigenic variation
- PMID: 25576484
- PMCID: PMC4346566
- DOI: 10.1128/EC.00207-14
DNA double-strand breaks and telomeres play important roles in trypanosoma brucei antigenic variation
Abstract
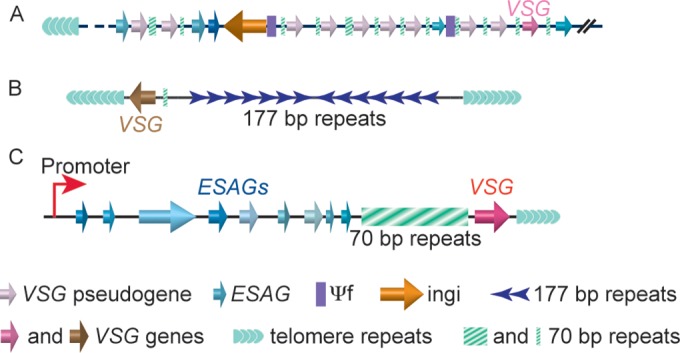
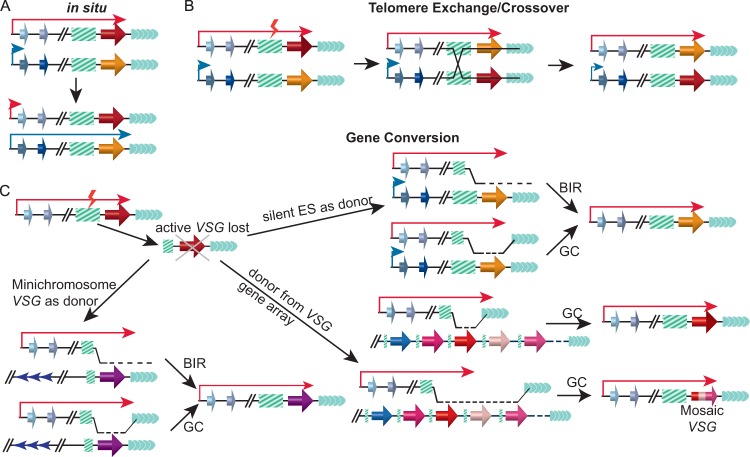
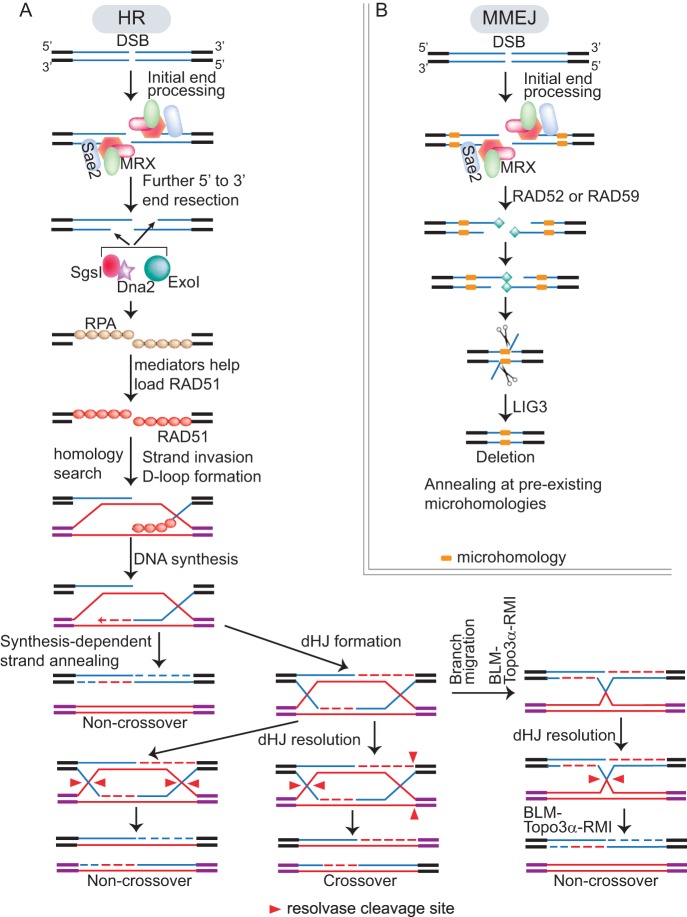
Human-infecting microbial pathogens all face a serious problem of elimination by the host immune response. Antigenic variation is an effective immune evasion mechanism where the pathogen regularly switches its major surface antigen. In many cases, the major surface antigen is encoded by genes from the same gene family, and its expression is strictly monoallelic. Among pathogens that undergo antigenic variation, Trypanosoma brucei (a kinetoplastid), which causes human African trypanosomiasis, Plasmodium falciparum (an apicomplexan), which causes malaria, Pneumocystis jirovecii (a fungus), which causes pneumonia, and Borrelia burgdorferi (a bacterium), which causes Lyme disease, also express their major surface antigens from loci next to the telomere. Except for Plasmodium, DNA recombination-mediated gene conversion is a major pathway for surface antigen switching in these pathogens. In the last decade, more sophisticated molecular and genetic tools have been developed in T. brucei, and our knowledge of functions of DNA recombination in antigenic variation has been greatly advanced. VSG is the major surface antigen in T. brucei. In subtelomeric VSG expression sites (ESs), VSG genes invariably are flanked by a long stretch of upstream 70-bp repeats. Recent studies have shown that DNA double-strand breaks (DSBs), particularly those in 70-bp repeats in the active ES, are a natural potent trigger for antigenic variation in T. brucei. In addition, telomere proteins can influence VSG switching by reducing the DSB amount at subtelomeric regions. These findings will be summarized and their implications will be discussed in this review.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Berriman M, Ghedin E, Hertz-Fowler C, Blandin G, Renauld H, Bartholomeu DC, Lennard NJ, Caler E, Hamlin NE, Haas B, Bohme U, Hannick L, Aslett MA, Shallom J, Marcello L, Hou L, Wickstead B, Alsmark UC, Arrowsmith C, Atkin RJ, Barron AJ, Bringaud F, Brooks K, Carrington M, Cherevach I, Chillingworth TJ, Churcher C, Clark LN, Corton CH, Cronin A, Davies RM, Doggett J, Djikeng A, Feldblyum T, Field MC, Fraser A, Goodhead I, Hance Z, Harper D, Harris BR, Hauser H, Hostetler J, Ivens A, Jagels K, Johnson D, Johnson J, Jones K, Kerhornou AX, Koo H, Larke N, Landfear S, Larkin C, Leech V, Line A, Lord A, Macleod A, Mooney PJ, Moule S, Martin DM, Morgan GW, Mungall K, Norbertczak H, Ormond D, Pai G, Peacock CS, Peterson J, Quail MA, Rabbinowitsch E, Rajandream MA, Reitter C, Salzberg SL, Sanders M, Schobel S, Sharp S, Simmonds M, Simpson AJ, Tallon L, Turner CM, Tait A, Tivey AR, Van Aken S, Walker D, Wanless D, Wang S, White B, White O, Whitehead S, Woodward J, Wortman J, Adams MD, Embley TM, Gull K, Ullu E, Barry JD, Fairlamb AH, Opperdoes F, Barrell BG, Donelson JE, Hall N, Fraser CM, Melville SE, El-Sayed NM. 2005. The genome of the African trypanosome Trypanosoma brucei. Science 309:416–422. doi: 10.1126/science.1112642. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
