

Nucleosome contact triggers conformational changes of Rpd3S driving high-affinity H3K36me nucleosome engagement
- PMID: 25578729
- PMCID: PMC4359074
- DOI: 10.1016/j.celrep.2014.12.027
Nucleosome contact triggers conformational changes of Rpd3S driving high-affinity H3K36me nucleosome engagement
Abstract
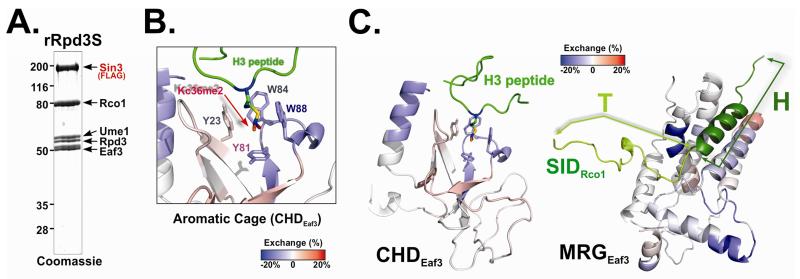
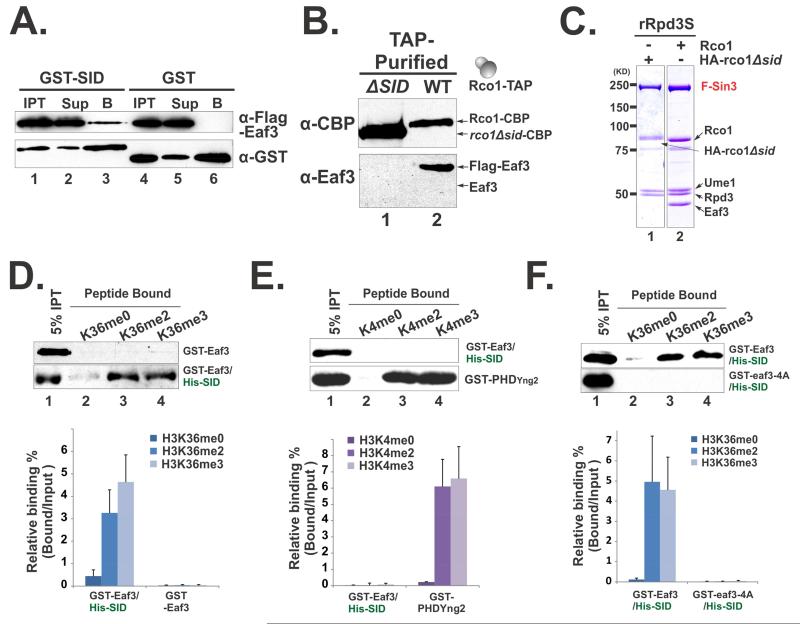
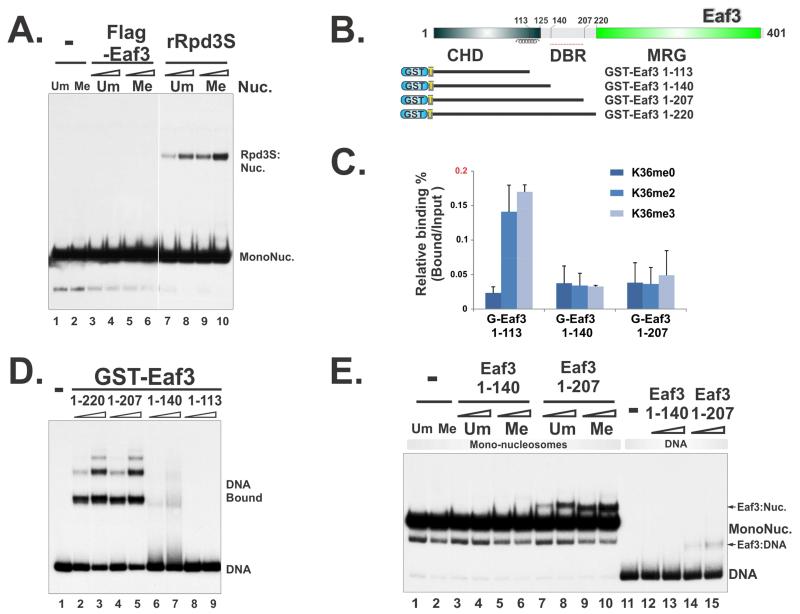
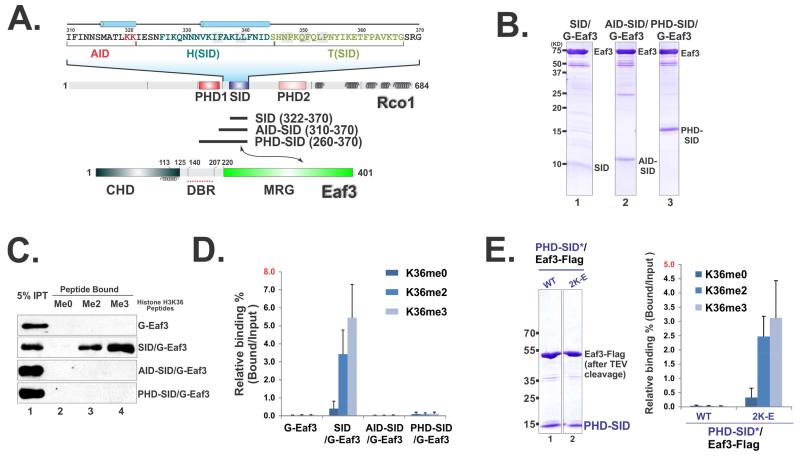
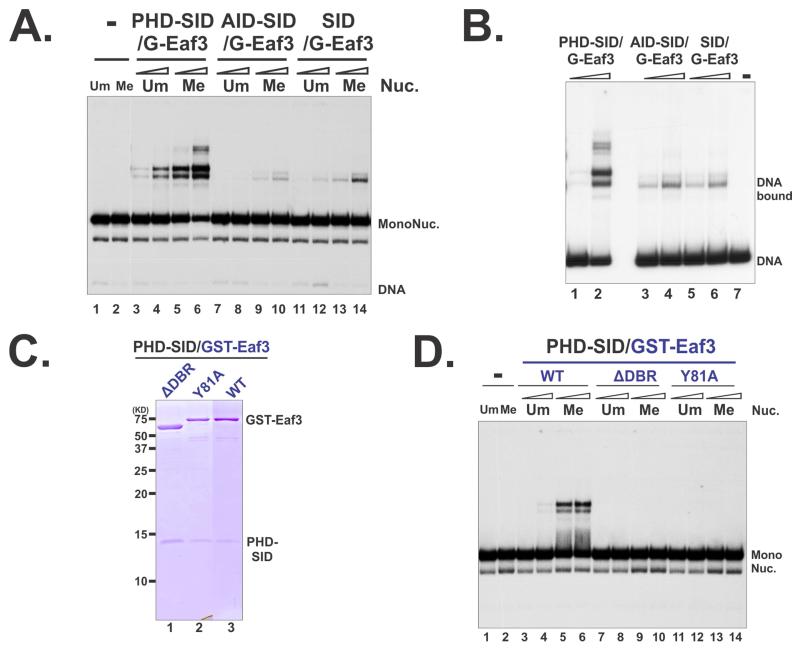
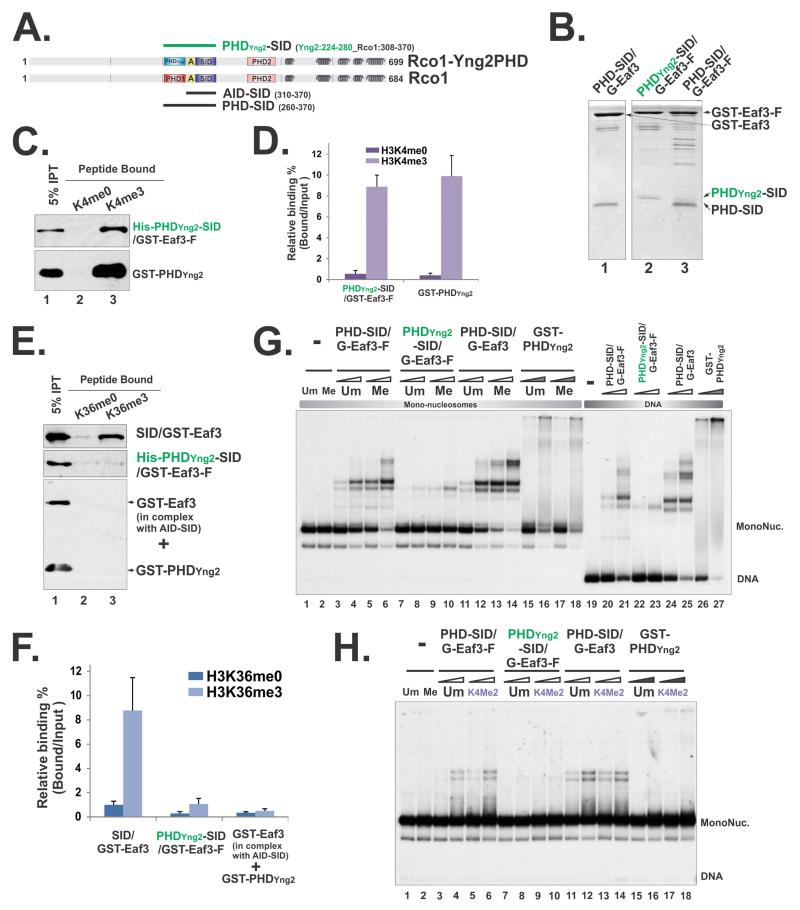
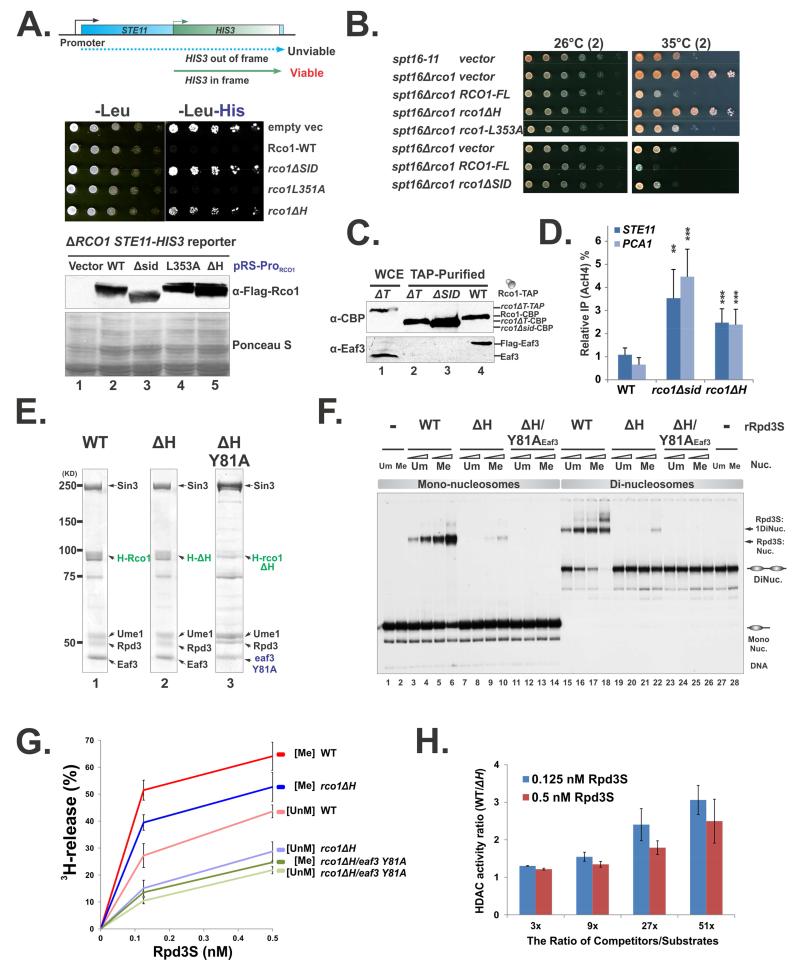
The Rpd3S histone deacetylase complex utilizes two subunits, Eaf3 and Rco1, to recognize nucleosomes methylated at H3K36 (H3K36me) with high affinity and strong specificity. However, the chromobarrel domain of Eaf3 (CHD) that is responsible for H3K36me recognition only binds weakly and with little specificity to histone peptides. Here, using deuterium exchange mass spectrometry (DXMS), we detected conformational changes of Rpd3S upon its contact with chromatin. Interestingly, we found that the Sin3-interacting domain of Rco1 (SID) allosterically stimulates preferential binding of Eaf3 to H3K36-methylated peptides. This activation is tightly regulated by an autoinhibitory mechanism to ensure optimal multivalent engagement of Rpd3S with nucleosomes. Lastly, we identified mutations at the interface between SID and Eaf3 that do not disrupt complex integrity but severely compromise Rpd3S functions in vitro and in vivo, suggesting that the nucleosome-induced conformational changes are essential for chromatin recognition.
Copyright © 2015 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Homodimeric PHD Domain-containing Rco1 Subunit Constitutes a Critical Interaction Hub within the Rpd3S Histone Deacetylase Complex.J Biol Chem. 2016 Mar 4;291(10):5428-38. doi: 10.1074/jbc.M115.703637. Epub 2016 Jan 8. J Biol Chem. 2016. PMID: 26747610 Free PMC article.
-
Combined action of PHD and chromo domains directs the Rpd3S HDAC to transcribed chromatin.Science. 2007 May 18;316(5827):1050-4. doi: 10.1126/science.1139004. Science. 2007. PMID: 17510366
-
Structure of histone deacetylase complex Rpd3S bound to nucleosome.Nat Struct Mol Biol. 2023 Dec;30(12):1893-1901. doi: 10.1038/s41594-023-01121-5. Epub 2023 Oct 5. Nat Struct Mol Biol. 2023. PMID: 37798513
-
Xenopus transcription factor IIIA and the 5S nucleosome: development of a useful in vitro system.Biochem Cell Biol. 2003 Jun;81(3):177-84. doi: 10.1139/o03-043. Biochem Cell Biol. 2003. PMID: 12897852 Review.
-
[Structures of proteins that regulate chromatin structures].Tanpakushitsu Kakusan Koso. 2000 Jun;45(9 Suppl):1658-70. Tanpakushitsu Kakusan Koso. 2000. PMID: 10879148 Review. Japanese. No abstract available.
Cited by
-
Structure and flexibility of the yeast NuA4 histone acetyltransferase complex.Elife. 2022 Oct 20;11:e81400. doi: 10.7554/eLife.81400. Elife. 2022. PMID: 36263929 Free PMC article.
-
The Eaf3 chromodomain acts as a pH sensor for gene expression by altering its binding affinity for histone methylated-lysine residues.Biosci Rep. 2020 Feb 28;40(2):BSR20191958. doi: 10.1042/BSR20191958. Biosci Rep. 2020. PMID: 32031206 Free PMC article.
-
Unique and Shared Roles for Histone H3K36 Methylation States in Transcription Regulation Functions.Cell Rep. 2020 Jun 9;31(10):107751. doi: 10.1016/j.celrep.2020.107751. Cell Rep. 2020. PMID: 32521276 Free PMC article.
-
Touch, act and go: landing and operating on nucleosomes.EMBO J. 2016 Feb 15;35(4):376-88. doi: 10.15252/embj.201593377. Epub 2016 Jan 19. EMBO J. 2016. PMID: 26787641 Free PMC article. Review.
-
Combined Action of Histone Reader Modules Regulates NuA4 Local Acetyltransferase Function but Not Its Recruitment on the Genome.Mol Cell Biol. 2016 Oct 28;36(22):2768-2781. doi: 10.1128/MCB.00112-16. Print 2016 Nov 15. Mol Cell Biol. 2016. PMID: 27550811 Free PMC article.
References
-
- Carrozza MJ, Li B, Florens L, Suganuma T, Swanson SK, Lee KK, Shia WJ, Anderson S, Yates J, Washburn MP, Workman JL. Histone H3 methylation by Set2 directs deacetylation of coding regions by Rpd3S to suppress spurious intragenic transcription. Cell. 2005;123:581–592. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
