

The energy landscape of adenylate kinase during catalysis
- PMID: 25580578
- PMCID: PMC4318763
- DOI: 10.1038/nsmb.2941
The energy landscape of adenylate kinase during catalysis
Abstract
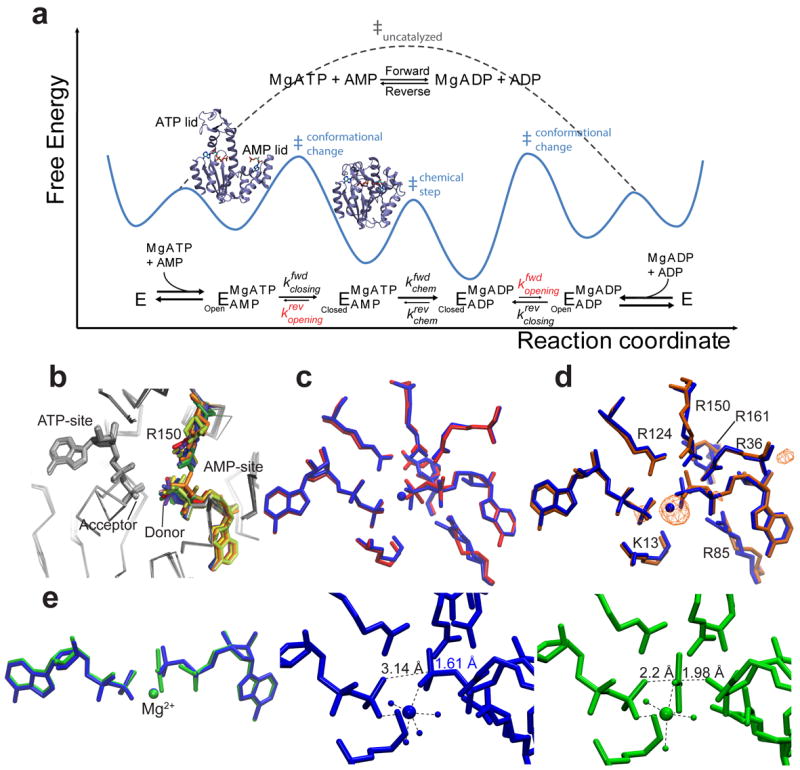
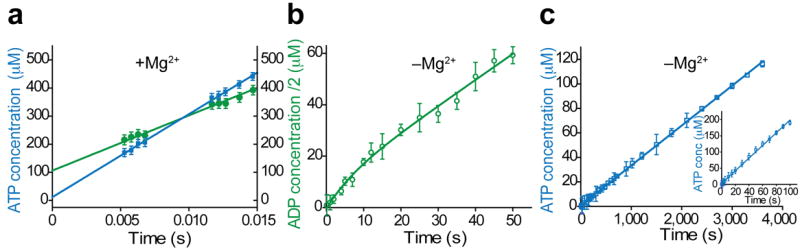
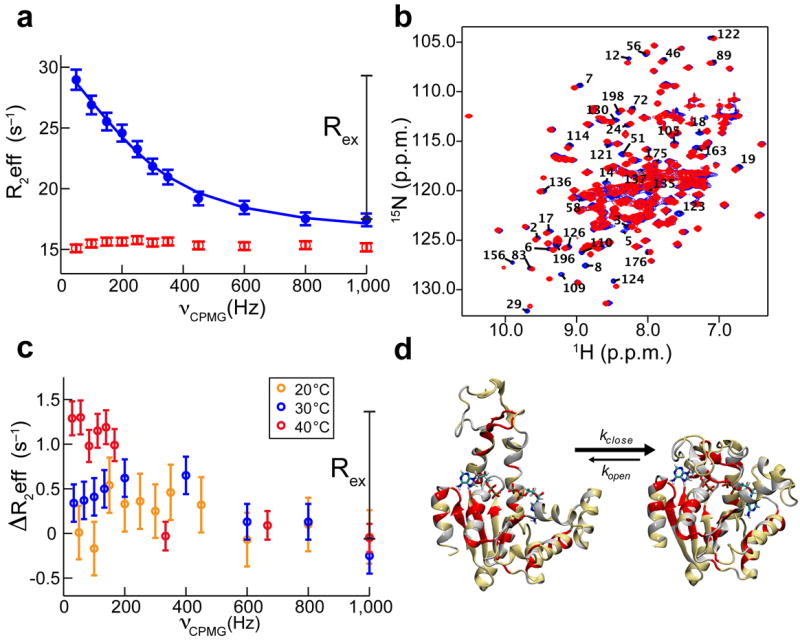
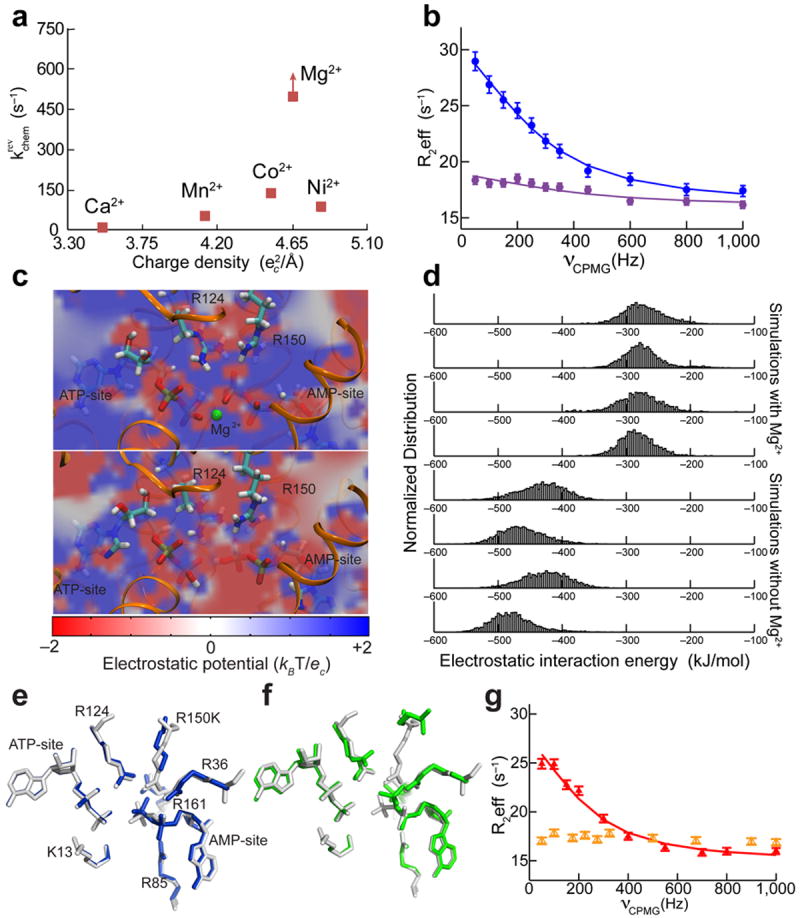
Kinases perform phosphoryl-transfer reactions in milliseconds; without enzymes, these reactions would take about 8,000 years under physiological conditions. Despite extensive studies, a comprehensive understanding of kinase energy landscapes, including both chemical and conformational steps, is lacking. Here we scrutinize the microscopic steps in the catalytic cycle of adenylate kinase, through a combination of NMR measurements during catalysis, pre-steady-state kinetics, molecular-dynamics simulations and crystallography of active complexes. We find that the Mg(2+) cofactor activates two distinct molecular events: phosphoryl transfer (>10(5)-fold) and lid opening (10(3)-fold). In contrast, mutation of an essential active site arginine decelerates phosphoryl transfer 10(3)-fold without substantially affecting lid opening. Our results highlight the importance of the entire energy landscape in catalysis and suggest that adenylate kinases have evolved to activate key processes simultaneously by precise placement of a single, charged and very abundant cofactor in a preorganized active site.
Figures

Comment in
-
An enzyme cofactor with a split personality.Nat Struct Mol Biol. 2015 Feb;22(2):101-3. doi: 10.1038/nsmb.2968. Nat Struct Mol Biol. 2015. PMID: 25650903 No abstract available.
References
-
- Westheimer FH. Why nature chose phosphates. Science. 1987;235:1173–1178. - PubMed
-
- Bowler MW, Cliff MJ, Waltho JP, Blackburn GM. Why did Nature select phosphate for its dominant roles in biology? New Journal of Chemistry. 2010;34:784–794.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
