

IL-33 facilitates oncogene-induced cholangiocarcinoma in mice by an interleukin-6-sensitive mechanism
- PMID: 25580681
- PMCID: PMC4406813
- DOI: 10.1002/hep.27687
IL-33 facilitates oncogene-induced cholangiocarcinoma in mice by an interleukin-6-sensitive mechanism
Abstract
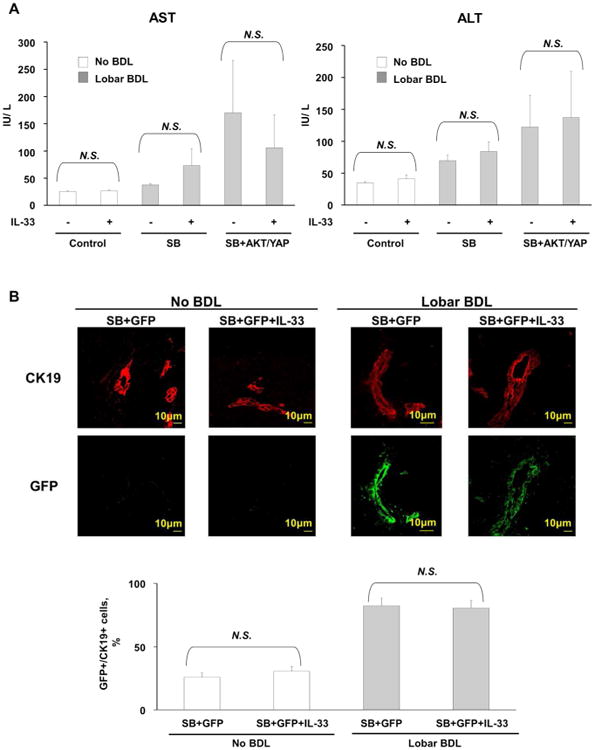
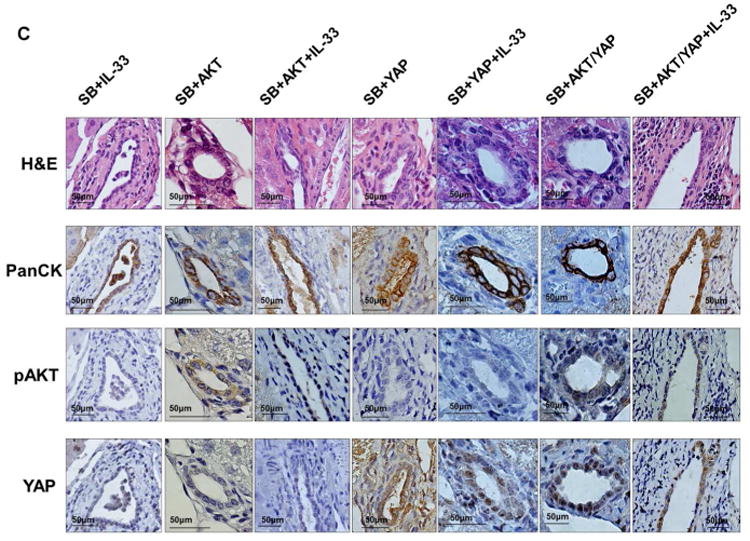
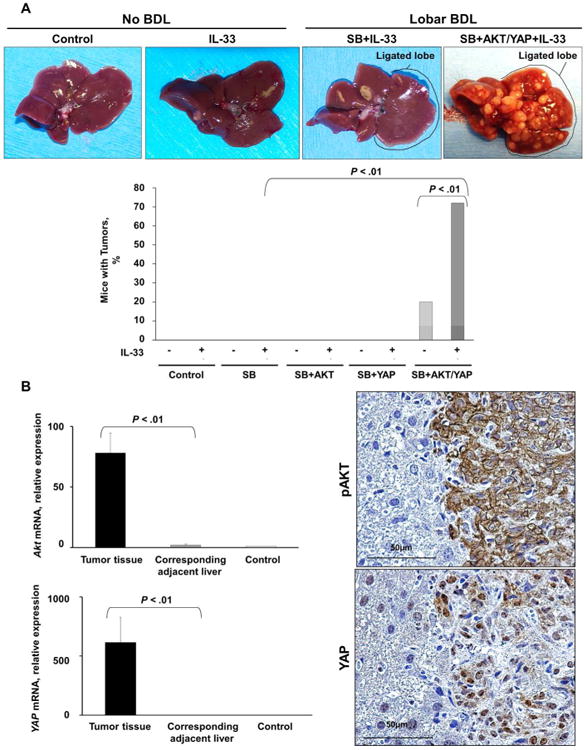
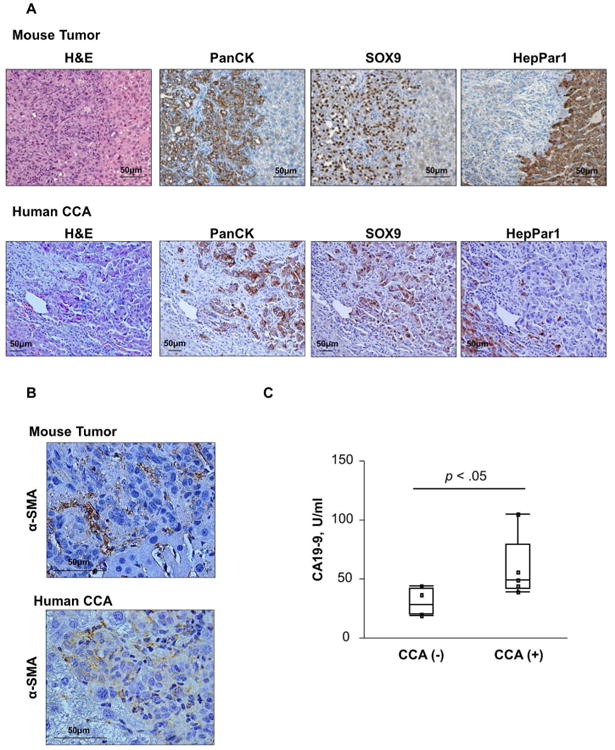
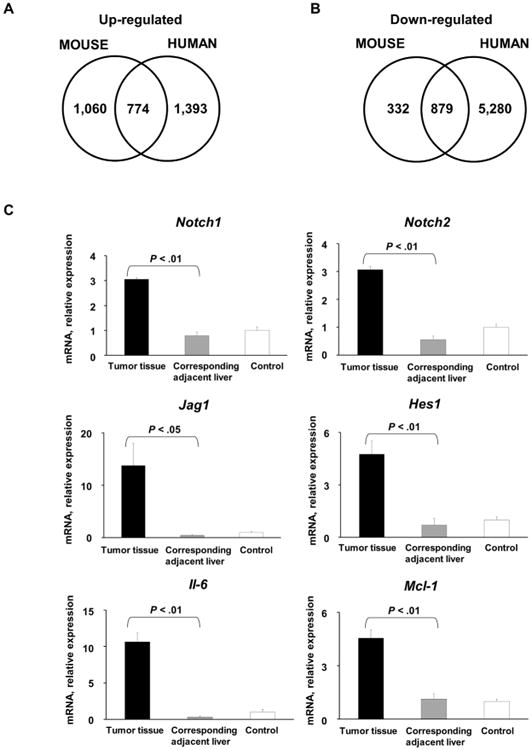
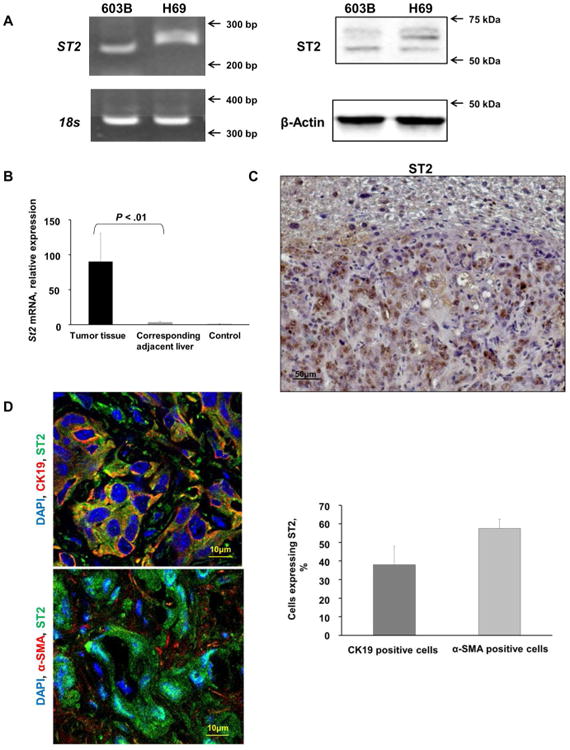
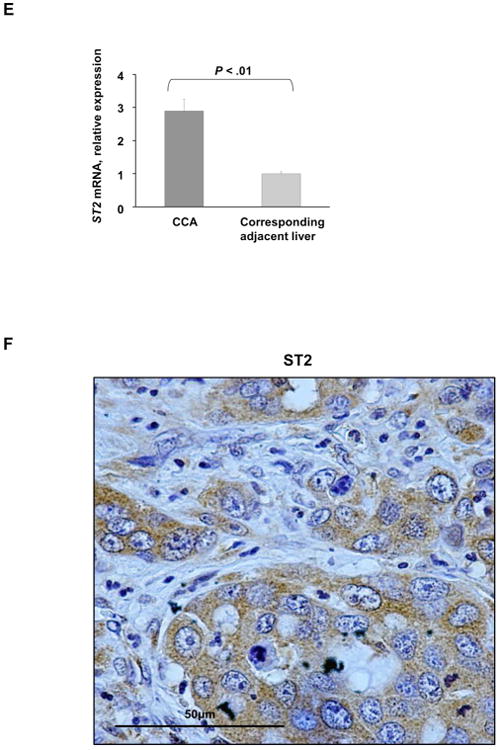
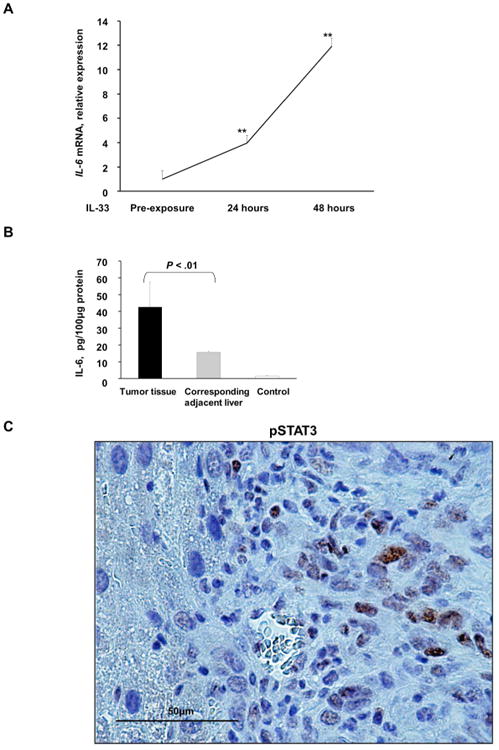
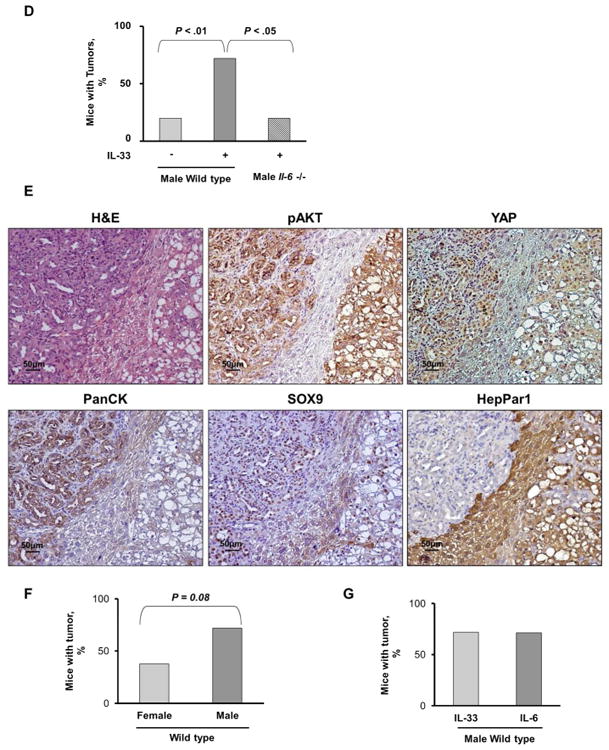
Cholangiocarcinoma (CCA) is a lethal hepatobiliary neoplasm originating from the biliary apparatus. In humans, CCA risk factors include hepatobiliary inflammation and fibrosis. The recently identified interleukin (IL)-1 family member, IL-33, has been shown to be a biliary mitogen which also promotes liver inflammation and fibrosis. Our aim was to generate a mouse model of CCA mimicking the human disease. Ectopic oncogene expression in the biliary tract was accomplished by the Sleeping Beauty transposon transfection system with transduction of constitutively active AKT (myr-AKT) and Yes-associated protein. Intrabiliary instillation of the transposon-transposase complex was coupled with lobar bile duct ligation in C57BL/6 mice, followed by administration of IL-33 for 3 consecutive days. Tumors developed in 72% of the male mice receiving both oncogenes plus IL-33 by 10 weeks but in only 20% of the male mice transduced with the oncogenes alone. Tumors expressed SOX9 and pancytokeratin (features of CCA) but were negative for HepPar1 (a marker of hepatocellular carcinoma). Substantive overlap with human CCA specimens was revealed by RNA profiling. Not only did IL-33 induce IL-6 expression by human cholangiocytes but it likely facilitated tumor development in vivo by an IL-6-sensitive process as tumor development was significantly attenuated in Il-6(-/-) male animals. Furthermore, tumor formation occurred at a similar rate when IL-6 was substituted for IL-33 in this model.
Conclusion: The transposase-mediated transduction of constitutively active AKT and Yes-associated protein in the biliary epithelium coupled with lobar obstruction and IL-33 administration results in the development of CCA with morphological and biochemical features of the human disease; this model highlights the role of inflammatory cytokines in CCA oncogenesis.
© 2015 by the American Association for the Study of Liver Diseases.
Figures
Comment in
-
Biliary tract. A new mouse model that closely resembles human cholangiocarcinoma.Nat Rev Gastroenterol Hepatol. 2015 Mar;12(3):122. doi: 10.1038/nrgastro.2015.10. Epub 2015 Jan 27. Nat Rev Gastroenterol Hepatol. 2015. PMID: 25623200 No abstract available.
References
-
- Sia D, Hoshida Y, Villanueva A, Roayaie S, Ferrer J, Tabak B, Peix J, Sole M, Tovar V, Alsinet C, Cornella H, Klotzle B, Fan JB, Cotsoglou C, Thung SN, Fuster J, Waxman S, Garcia-Valdecasas JC, Bruix J, Schwartz ME, Beroukhim R, Mazzaferro V, Llovet JM. Integrative molecular analysis of intrahepatic cholangiocarcinoma reveals 2 classes that have different outcomes. Gastroenterology. 2013;144:829–40. - PMC - PubMed
-
- Sirica AE. The role of cancer-associated myofibroblasts in intrahepatic cholangiocarcinoma. Nat Rev Gastroenterol Hepatol. 2012;9:44–54. - PubMed
-
- Everhart JE, Ruhl CE. Burden of digestive diseases in the United States Part III: Liver, biliary tract, and pancreas. Gastroenterology. 2009;136:1134–44. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01CA136606/CA/NCI NIH HHS/United States
- DK64008/DK/NIDDK NIH HHS/United States
- P30 DK026743/DK/NIDDK NIH HHS/United States
- T32 DK007198/DK/NIDDK NIH HHS/United States
- K12 CA90628/CA/NCI NIH HHS/United States
- R01 DK059427/DK/NIDDK NIH HHS/United States
- R56 DK059427/DK/NIDDK NIH HHS/United States
- K12 CA090628/CA/NCI NIH HHS/United States
- T32DK007198/DK/NIDDK NIH HHS/United States
- DK84567/DK/NIDDK NIH HHS/United States
- P30 CA015083/CA/NCI NIH HHS/United States
- R01 CA136606/CA/NCI NIH HHS/United States
- DK59427/DK/NIDDK NIH HHS/United States
- P30 DK084567/DK/NIDDK NIH HHS/United States
- R01 DK064008/DK/NIDDK NIH HHS/United States
- DK83781/DK/NIDDK NIH HHS/United States
- R01 DK083781/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials