

Competition with and without priority control: linking rivalry to attention through winner-take-all networks with memory
- PMID: 25581077
- PMCID: PMC4376592
- DOI: 10.1111/nyas.12575
Competition with and without priority control: linking rivalry to attention through winner-take-all networks with memory
Abstract
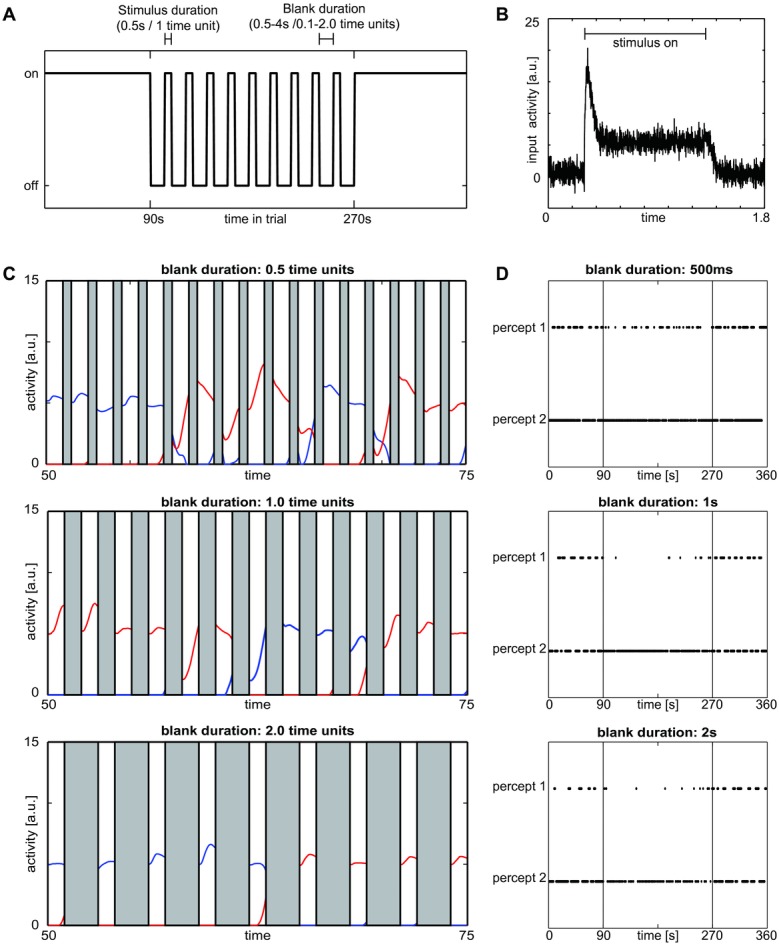
Competition is ubiquitous in perception. For example, items in the visual field compete for processing resources, and attention controls their priority (biased competition). The inevitable ambiguity in the interpretation of sensory signals yields another form of competition: distinct perceptual interpretations compete for access to awareness. Rivalry, where two equally likely percepts compete for dominance, explicates the latter form of competition. Building upon the similarity between attention and rivalry, we propose to model rivalry by a generic competitive circuit that is widely used in the attention literature-a winner-take-all (WTA) network. Specifically, we show that a network of two coupled WTA circuits replicates three common hallmarks of rivalry: the distribution of dominance durations, their dependence on input strength ("Levelt's propositions"), and the effects of stimulus removal (blanking). This model introduces a form of memory by forming discrete states and explains experimental data better than competitive models of rivalry without memory. This result supports the crucial role of memory in rivalry specifically and in competitive processes in general. Our approach unifies the seemingly distinct phenomena of rivalry, memory, and attention in a single model with competition as the common underlying principle.
Keywords: attention; binocular rivalry; modeling; psychophysics; vision; winner-take-all network.
© 2015 The Authors. Annals of the New York Academy of Sciences published by Wiley Periodicals Inc. on behalf of The New York Academy of Sciences.
Figures





References
-
- James W. The Principles of Psychology. New York: Holt; 1890.
-
- Desimone R, Duncan J. Neural mechanisms of selective visual attention. Annu. Rev. Neurosci. 1995;18:193–222. - PubMed
-
- Duncan J. “Cooperating brain systems in selective perception and action”. In: Inui T, McClelland JL, editors. Attention and Performance XVI: Information Integration in Perception and Communication. Cambridge, MA: MIT Press; 1996. pp. 549–578.
-
- Duncan J. EPS Mid-Career Award 2004: brain mechanisms of attention. Q. J. Exp. Psychol. 2006;59:2–27. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
