

Autophagy-independent functions of UVRAG are essential for peripheral naive T-cell homeostasis
- PMID: 25583492
- PMCID: PMC4313859
- DOI: 10.1073/pnas.1423588112
Autophagy-independent functions of UVRAG are essential for peripheral naive T-cell homeostasis
Abstract
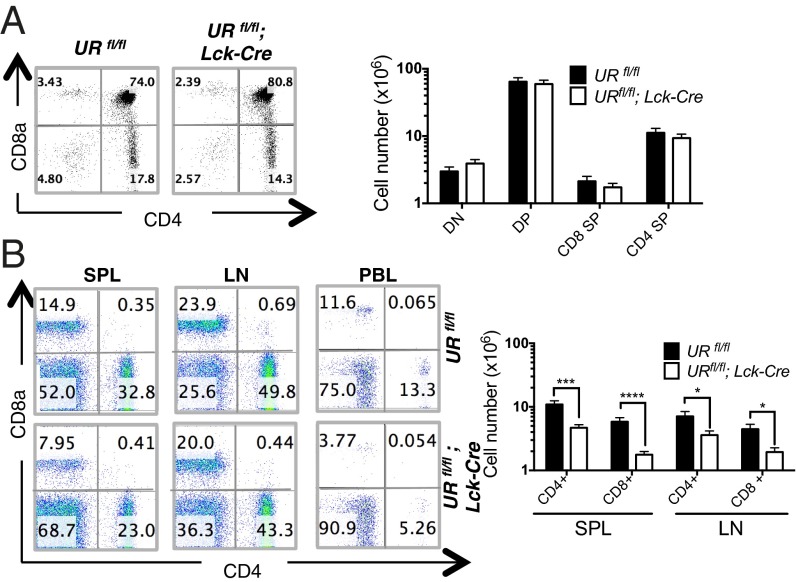
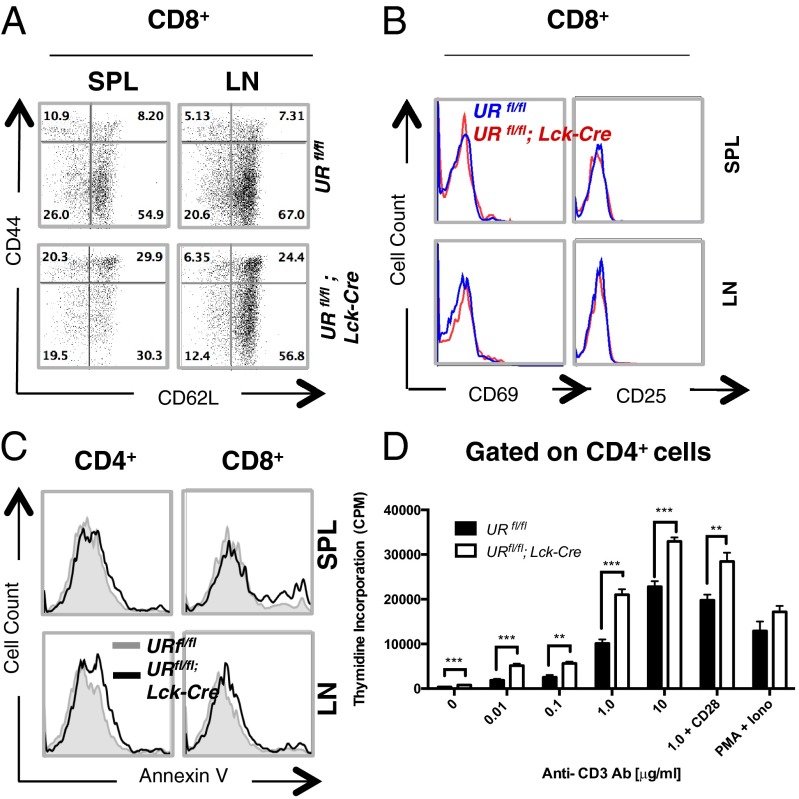
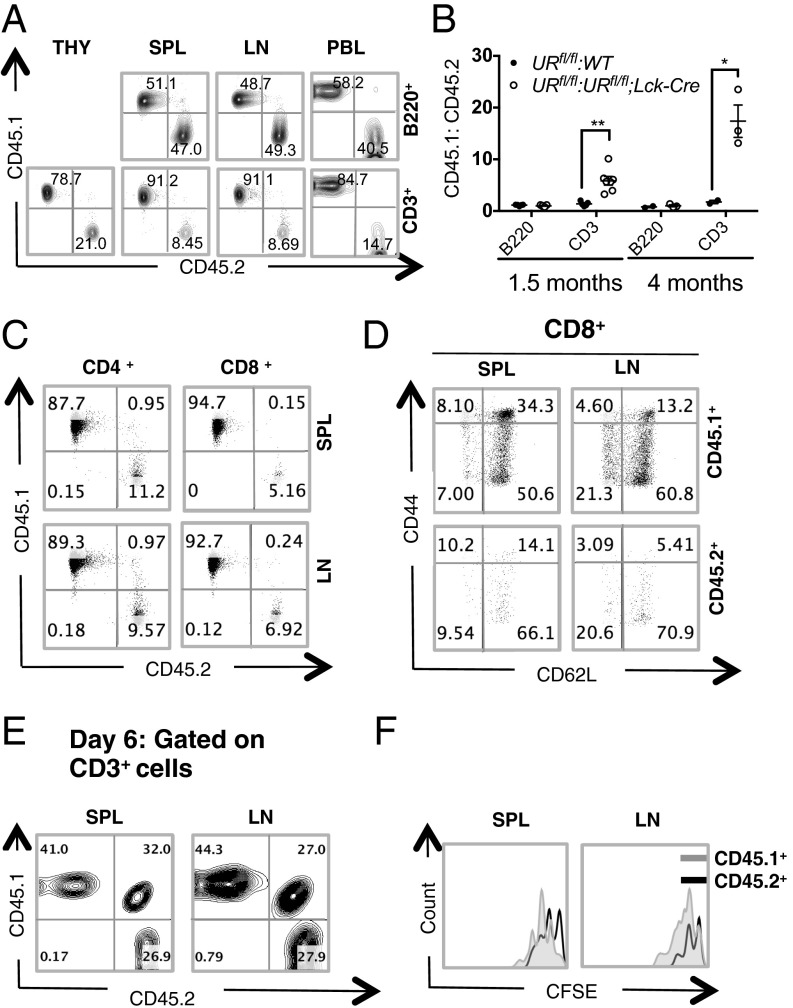
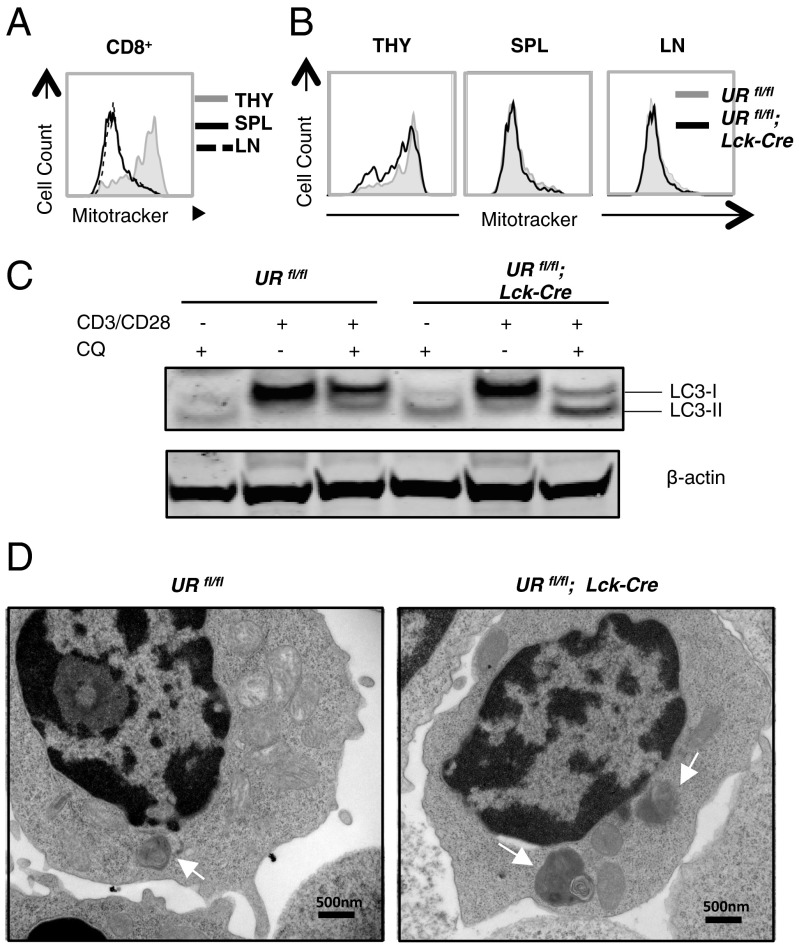
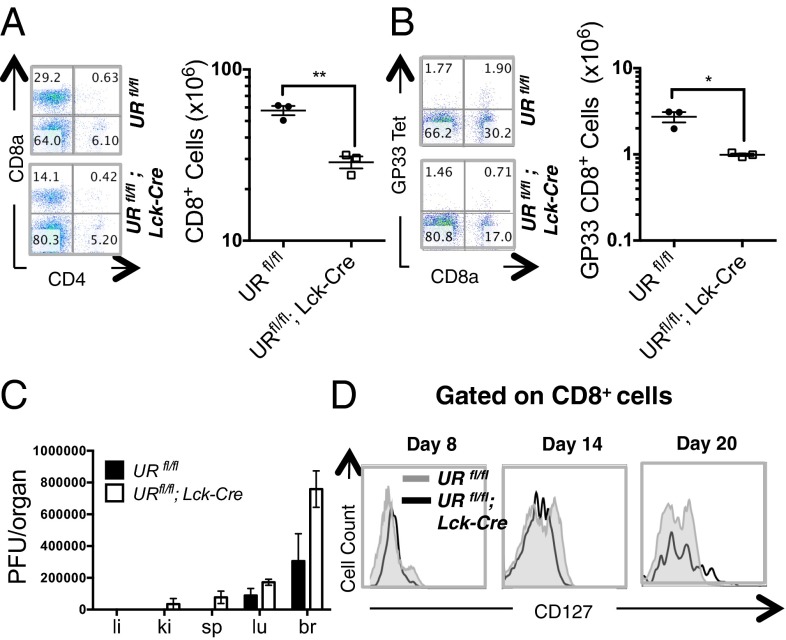
UV radiation resistance-associated gene (UVRAG) encodes a tumor suppressor with putative roles in autophagy, endocytic trafficking, and DNA damage repair but its in vivo role in T cells is unknown. Because conditional homozygous deletion of Uvrag in mice results in early embryonic lethality, we generated T-cell-specific UVRAG-deficient mice that lacked UVRAG expression specifically in T cells. This loss of UVRAG led to defects in peripheral homeostasis that could not be explained by the increased sensitivity to cell death and impaired proliferation observed for other autophagy-related gene knockout mice. Instead, UVRAG-deficient T-cells exhibited normal mitochondrial clearance and activation-induced autophagy, suggesting that UVRAG has an autophagy-independent role that is critical for peripheral naive T-cell homeostatic proliferation. In vivo, T-cell-specific loss of UVRAG dampened CD8(+) T-cell responses to LCMV infection in mice, delayed viral clearance, and impaired memory T-cell generation. Our data provide novel insights into the control of autophagy in T cells and identify UVRAG as a new regulator of naïve peripheral T-cell homeostasis.
Keywords: T-cell homeostasis; UVRAG-deficient mice; autophagy; embryonic lethality.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Li C, et al. Autophagy is induced in CD4+ T cells and important for the growth factor-withdrawal cell death. J Immunol. 2006;177(8):5163–5168. - PubMed
-
- Jia W, He YW. Temporal regulation of intracellular organelle homeostasis in T lymphocytes by autophagy. J Immunol. 2011;186(9):5313–5322. - PubMed
-
- Pua HH, Guo J, Komatsu M, He YW. Autophagy is essential for mitochondrial clearance in mature T lymphocytes. J Immunol. 2009;182(7):4046–4055. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
