

Translation initiation mediated by RNA looping
- PMID: 25583496
- PMCID: PMC4313796
- DOI: 10.1073/pnas.1416883112
Translation initiation mediated by RNA looping
Abstract
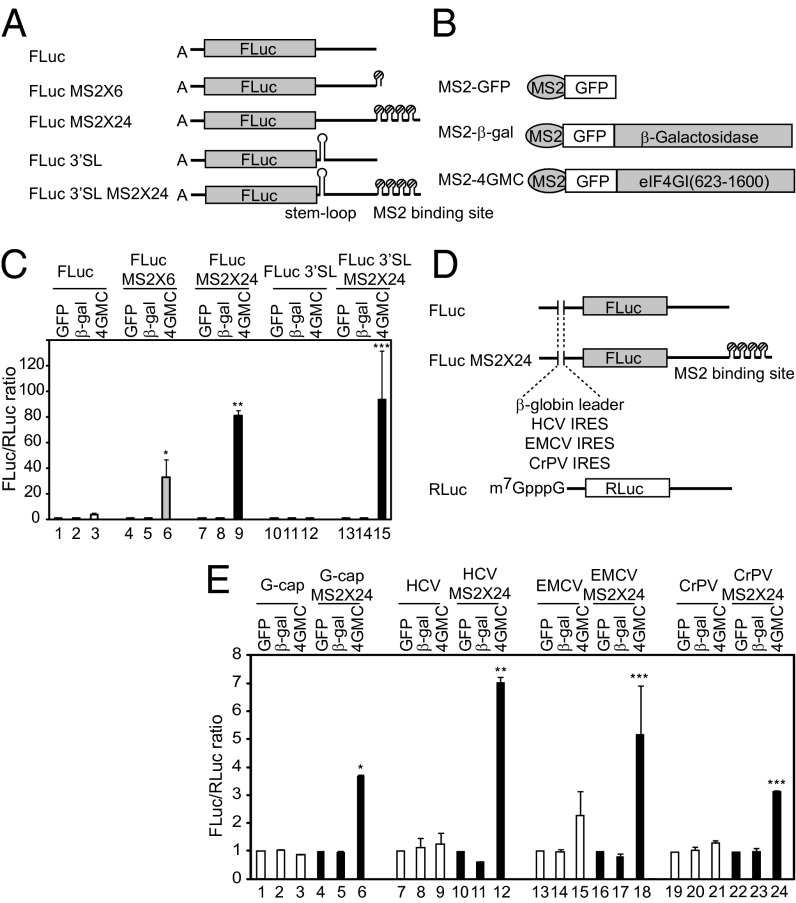
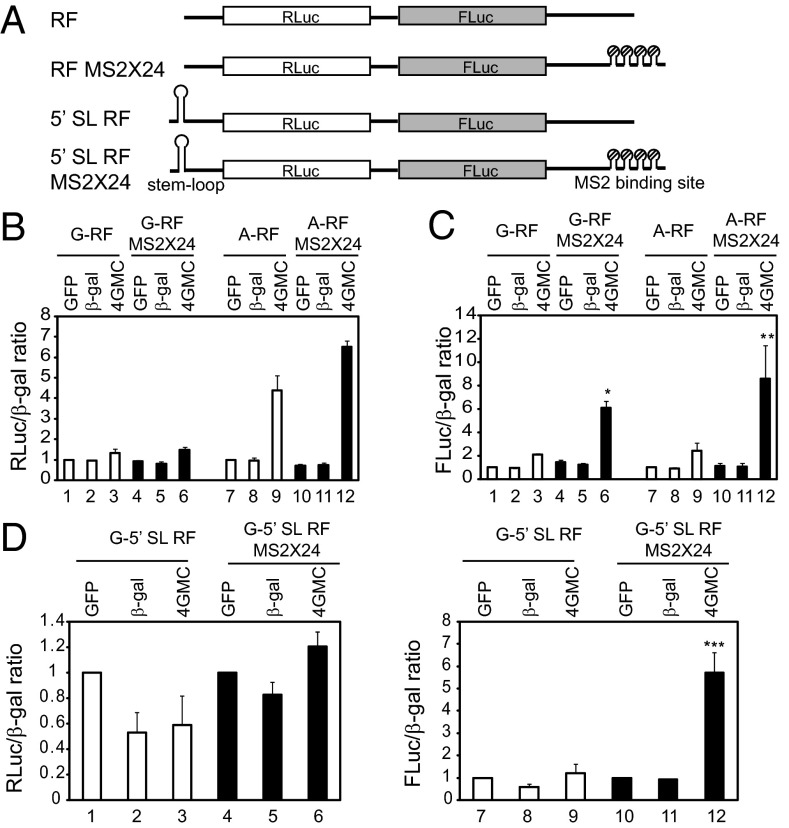
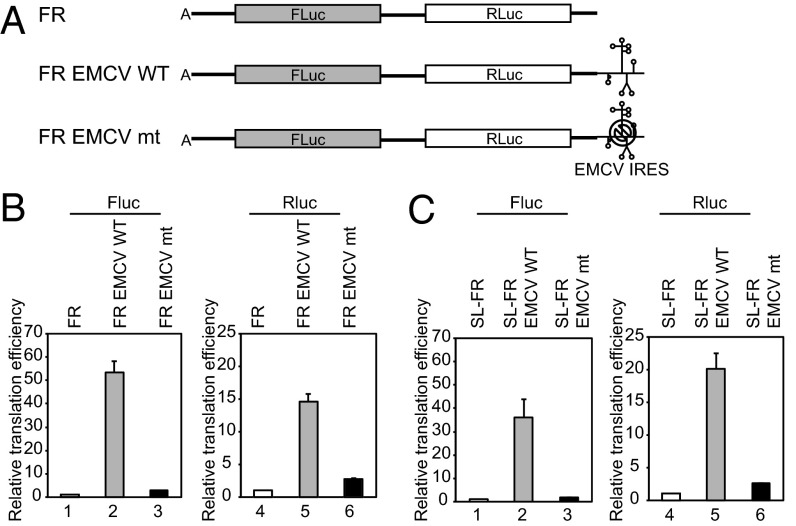
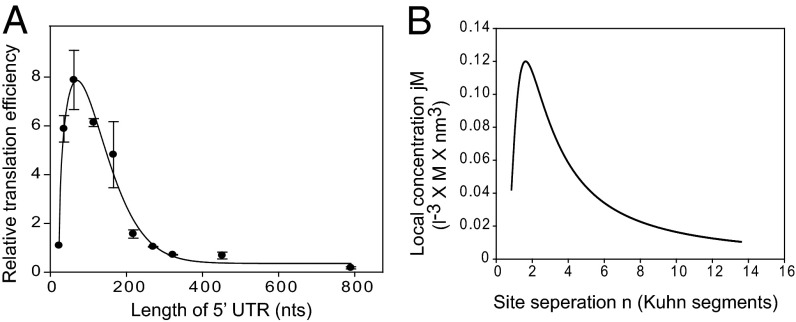
Eukaryotic translation initiation commences at the initiation codon near the 5' end of mRNA by a 40S ribosomal subunit, and the recruitment of a 40S ribosome to an mRNA is facilitated by translation initiation factors interacting with the m(7)G cap and/or poly(A) tail. The 40S ribosome recruited to an mRNA is then transferred to the AUG initiation codon with the help of translation initiation factors. To understand the mechanism by which the ribosome finds an initiation codon, we investigated the role of eIF4G in finding the translational initiation codon. An artificial polypeptide eIF4G fused with MS2 was localized downstream of the reporter gene through MS2-binding sites inserted in the 3' UTR of the mRNA. Translation of the reporter was greatly enhanced by the eIF4G-MS2 fusion protein regardless of the presence of a cap structure. Moreover, eIF4G-MS2 tethered at the 3' UTR enhanced translation of the second cistron of a dicistronic mRNA. The encephalomyocarditis virus internal ribosome entry site, a natural translational-enhancing element facilitating translation through an interaction with eIF4G, positioned downstream of a reporter gene, also enhanced translation of the upstream gene in a cap-independent manner. Finally, we mathematically modeled the effect of distance between the cap structure and initiation codon on the translation efficiency of mRNAs. The most plausible explanation for translational enhancement by the translational-enhancing sites is recognition of the initiation codon by the ribosome bound to the ribosome-recruiting sites through "RNA looping." The RNA looping hypothesis provides a logical explanation for augmentation of translation by enhancing elements located upstream and/or downstream of a protein-coding region.
Keywords: RNA looping; eukaryotic mRNA; ribosome scanning; translation initiation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Jackson RJ. Alternative mechanisms of initiating translation of mammalian mRNAs. Biochem Soc Trans. 2005;33(Pt 6):1231–1241. - PubMed
-
- Pelletier J, Sonenberg N. Internal initiation of translation of eukaryotic mRNA directed by a sequence derived from poliovirus RNA. Nature. 1988;334(6180):320–325. - PubMed
-
- Kolupaeva VG, Pestova TV, Hellen CU, Shatsky IN. Translation eukaryotic initiation factor 4G recognizes a specific structural element within the internal ribosome entry site of encephalomyocarditis virus RNA. J Biol Chem. 1998;273(29):18599–18604. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
