

The histone methyltransferase Smyd2 is a negative regulator of macrophage activation by suppressing interleukin 6 (IL-6) and tumor necrosis factor α (TNF-α) production
- PMID: 25583990
- PMCID: PMC4342458
- DOI: 10.1074/jbc.M114.610345
The histone methyltransferase Smyd2 is a negative regulator of macrophage activation by suppressing interleukin 6 (IL-6) and tumor necrosis factor α (TNF-α) production
Abstract
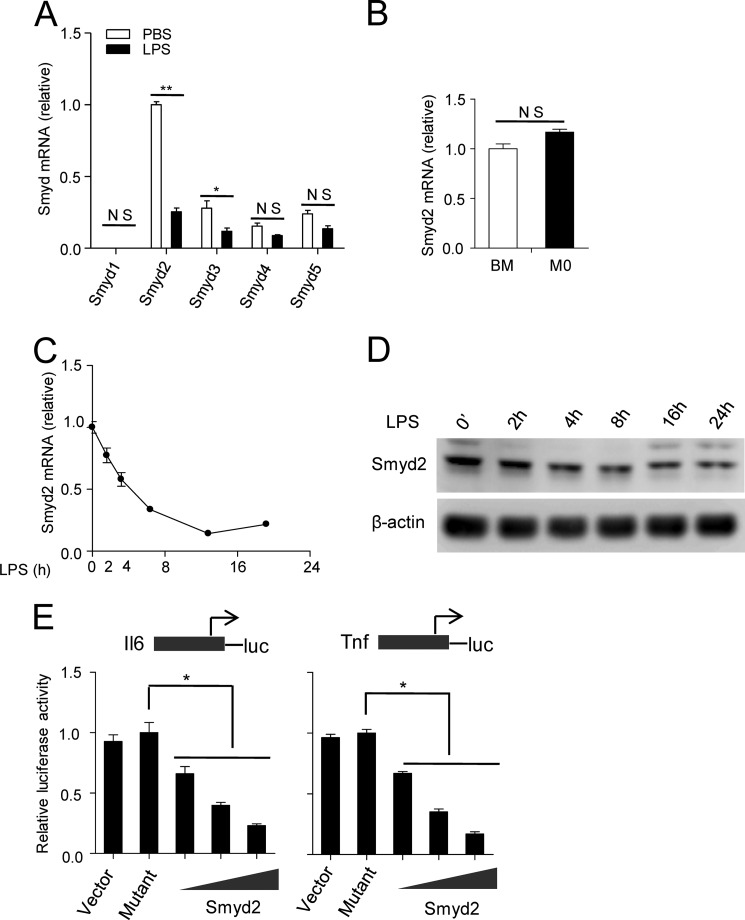
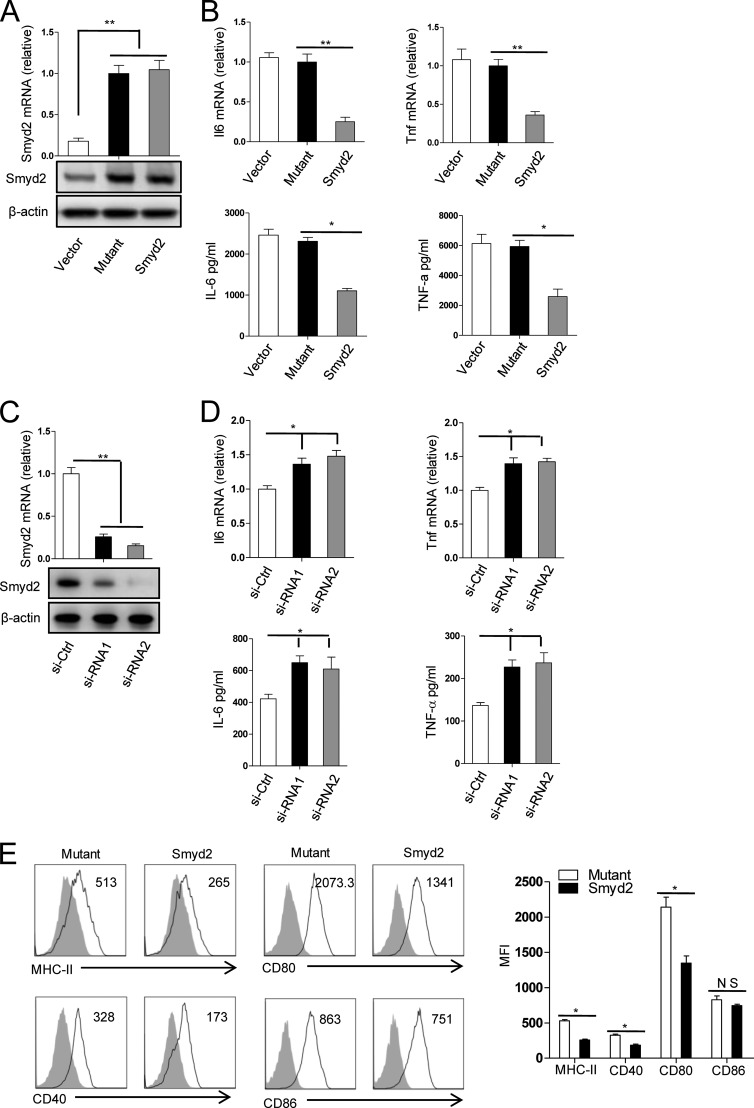
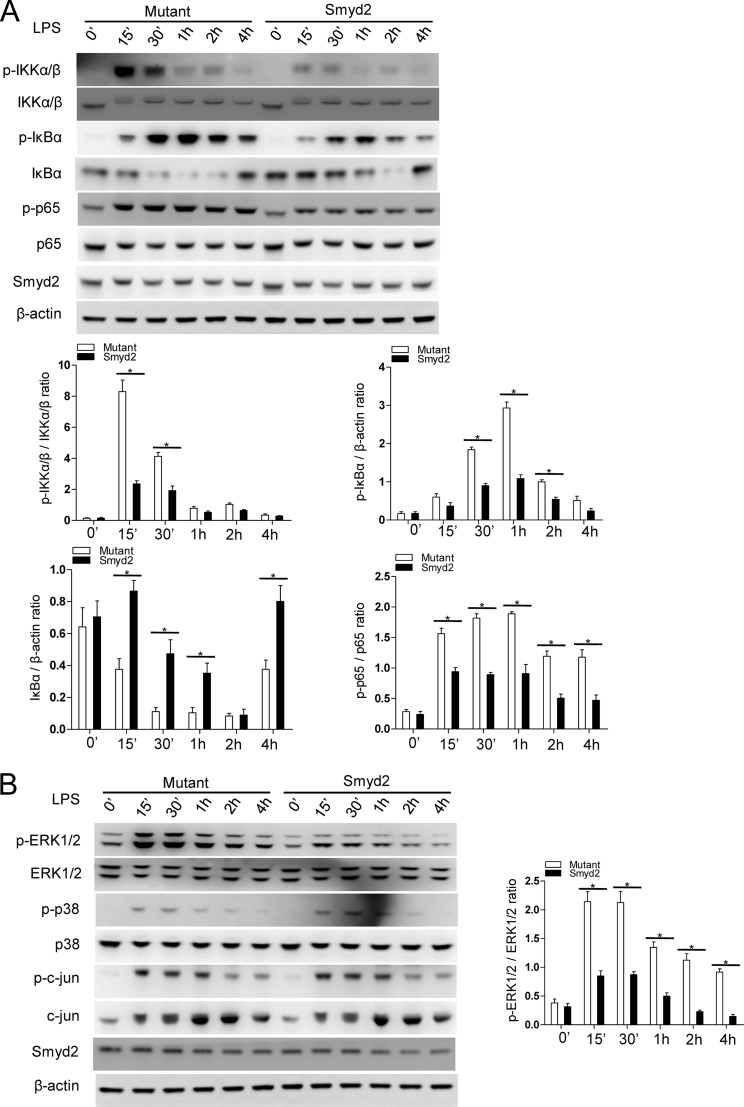
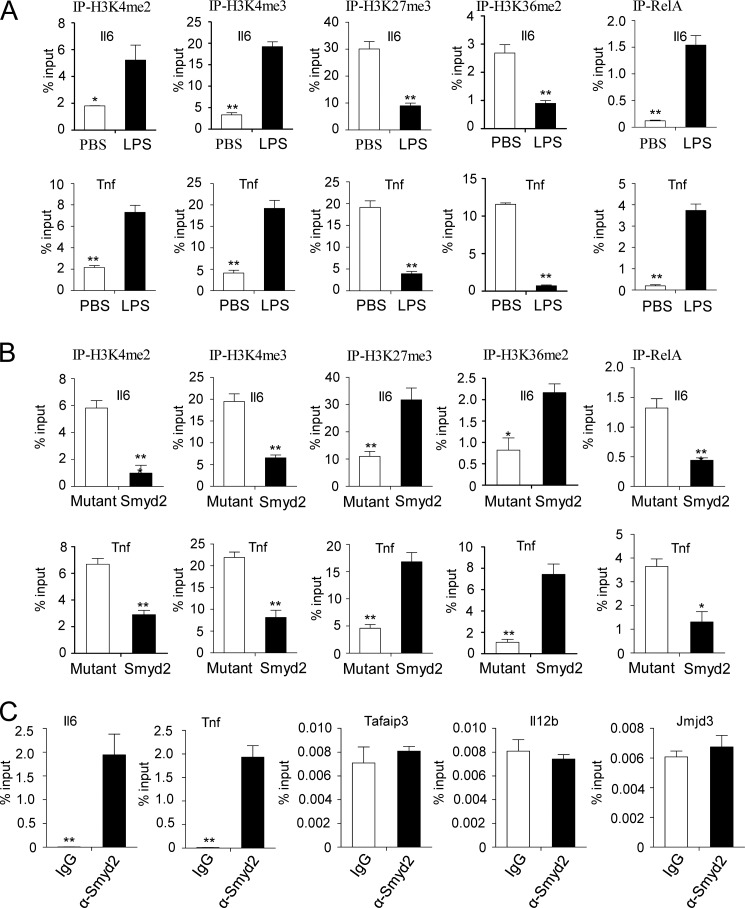
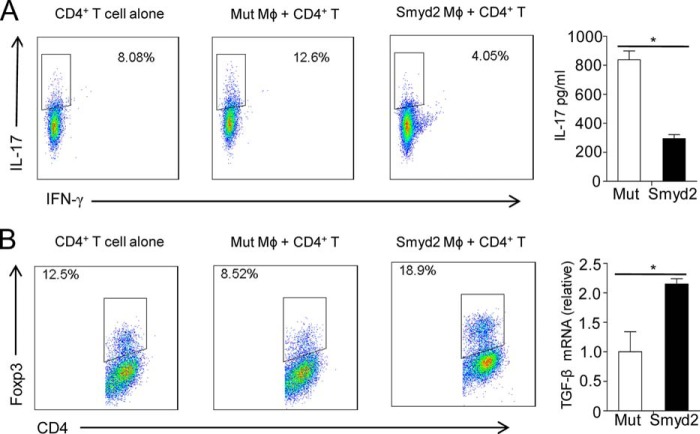
SET and MYND domain-containing 2 (Smyd2), a histone 3 lysine 4- and histone 3 lysine 36 (H3K36)-specific methyltransferase, plays critical roles in cardiac development and tumorigenesis. However, the role of Smyd2 in immunity and inflammation remains poorly understood. In this study, we report that Smyd2 is a novel negative regulator for macrophage activation and M1 polarization. Elevated Smyd2 expression suppresses the production of proinflammatory cytokines, including IL-6 and TNF, and inhibits the expression of important cell surface molecules, including major MHC-II and costimulatory molecules. Furthermore, macrophages with high Smyd2 expression inhibit Th-17 cell differentiation but promote regulatory T cell differentiation as a result of increased TGF-β production and decreased IL-6 secretion. In macrophages, Smyd2 specifically facilitates H3K36 dimethylation at Tnf and Il6 promoters to suppress their transcription and inhibits NF-κB and ERK signaling. Therefore, our data demonstrate that epigenetic modification by Smyd2-mediated H3K36 dimethylation at Tnf and Il6 promoters plays an important role in the regulation of macrophage activation during inflammation.
Keywords: Autoimmunity; ChiP; Immunology; Macrophage; Signaling.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
Similar articles
-
Regulation of estrogen receptor α by histone methyltransferase SMYD2-mediated protein methylation.Proc Natl Acad Sci U S A. 2013 Oct 22;110(43):17284-9. doi: 10.1073/pnas.1307959110. Epub 2013 Oct 7. Proc Natl Acad Sci U S A. 2013. PMID: 24101509 Free PMC article.
-
Leishmania donovani Subverts Host Immune Response by Epigenetic Reprogramming of Macrophage M(Lipopolysaccharides + IFN-γ)/M(IL-10) Polarization.J Immunol. 2020 May 15;204(10):2762-2778. doi: 10.4049/jimmunol.1900251. Epub 2020 Apr 10. J Immunol. 2020. PMID: 32277055
-
The H3K9 methyltransferase Setdb1 regulates TLR4-mediated inflammatory responses in macrophages.Sci Rep. 2016 Jun 28;6:28845. doi: 10.1038/srep28845. Sci Rep. 2016. PMID: 27349785 Free PMC article.
-
Histone methyltransferase SMYD2: ubiquitous regulator of disease.Clin Epigenetics. 2019 Aug 1;11(1):112. doi: 10.1186/s13148-019-0711-4. Clin Epigenetics. 2019. PMID: 31370883 Free PMC article. Review.
-
Regulation of the MIR155 host gene in physiological and pathological processes.Gene. 2013 Dec 10;532(1):1-12. doi: 10.1016/j.gene.2012.12.009. Epub 2012 Dec 14. Gene. 2013. PMID: 23246696 Review.
Cited by
-
EZH1 Is Associated with TCP-Induced Bone Regeneration through Macrophage Polarization.Stem Cells Int. 2018 Aug 30;2018:6310560. doi: 10.1155/2018/6310560. eCollection 2018. Stem Cells Int. 2018. PMID: 30228822 Free PMC article.
-
Inhibitors of Protein Methyltransferases and Demethylases.Chem Rev. 2018 Feb 14;118(3):989-1068. doi: 10.1021/acs.chemrev.6b00801. Epub 2017 Mar 24. Chem Rev. 2018. PMID: 28338320 Free PMC article. Review.
-
Epigenetic Regulation of Monocyte and Macrophage Function.Antioxid Redox Signal. 2016 Nov 10;25(14):758-774. doi: 10.1089/ars.2016.6695. Epub 2016 Apr 25. Antioxid Redox Signal. 2016. PMID: 26983461 Free PMC article. Review.
-
Spatial confinement downsizes the inflammatory response of macrophages.Nat Mater. 2018 Dec;17(12):1134-1144. doi: 10.1038/s41563-018-0190-6. Epub 2018 Oct 22. Nat Mater. 2018. PMID: 30349032 Free PMC article.
-
Bacterial Pathogens Hijack the Innate Immune Response by Activation of the Reverse Transsulfuration Pathway.mBio. 2019 Oct 29;10(5):e02174-19. doi: 10.1128/mBio.02174-19. mBio. 2019. PMID: 31662455 Free PMC article.
References
-
- Kugelberg E. (2014) Innate immunity: a wee protection. Nat. Rev. Immunol. 14, 359. - PubMed
-
- Bordon Y. (2012) Innate immunity: bitter enemies. Nat. Rev. Immunol. 12, 746. - PubMed
-
- Papatriantafyllou M. (2012) Innate immunity: inflammasome triggered by cell swelling. Nat. Rev. Immunol. 12, 742. - PubMed
-
- Takeda K., Kaisho T., Akira S. (2003) Toll-like receptors. Annu. Rev. Immunol. 21, 335–376 - PubMed
-
- O'Neill L. A., Golenbock D., Bowie A. G. (2013) The history of Toll-like receptors: redefining innate immunity. Nat. Rev. Immunol. 13, 453–460 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
