

Proteomic characterization of the nucleolar linker histone H1 interaction network
- PMID: 25584861
- PMCID: PMC4417401
- DOI: 10.1016/j.jmb.2015.01.001
Proteomic characterization of the nucleolar linker histone H1 interaction network
Abstract
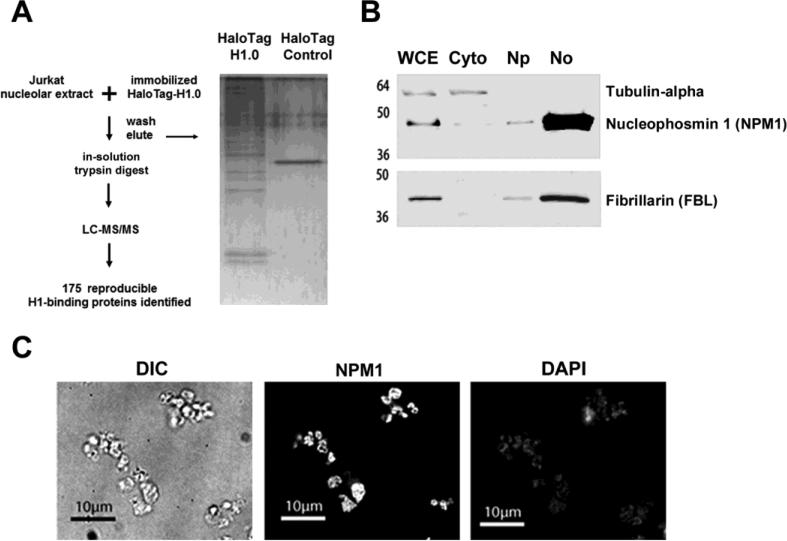
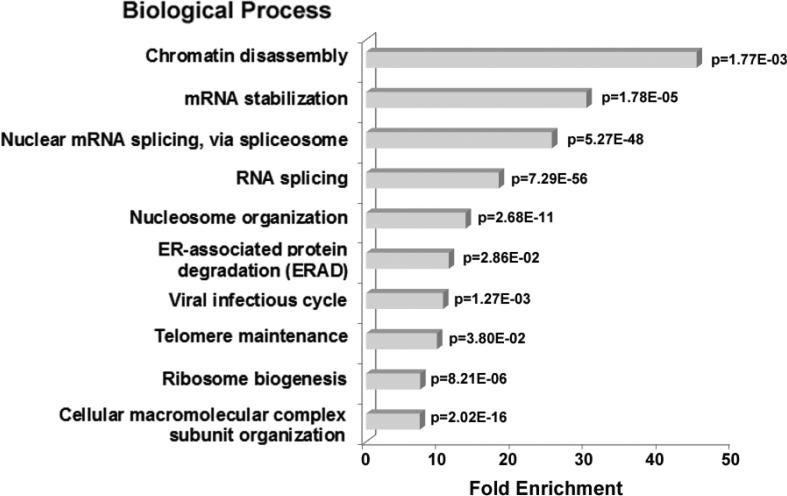
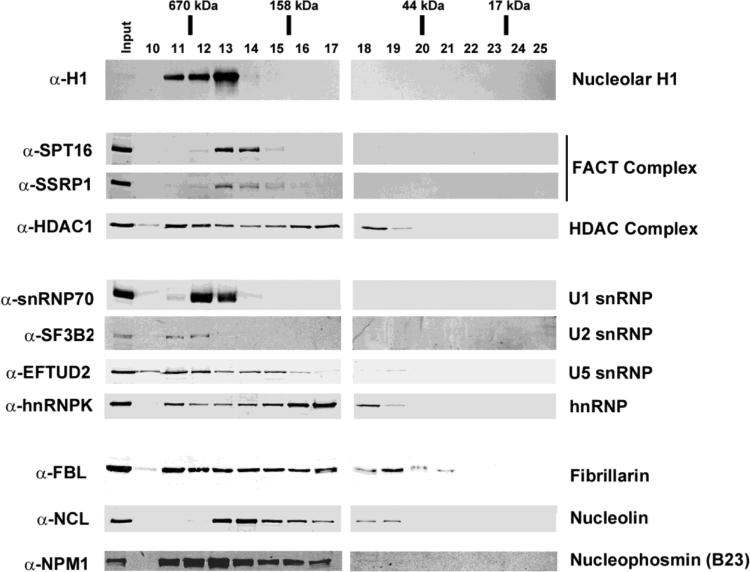
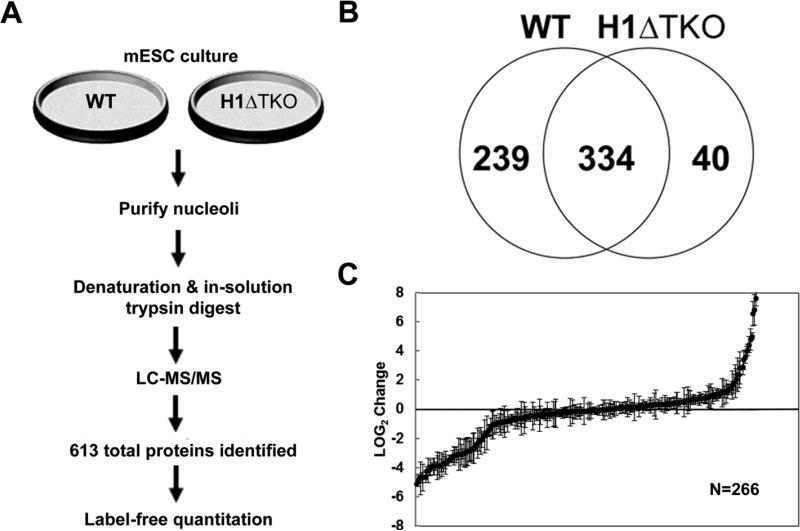
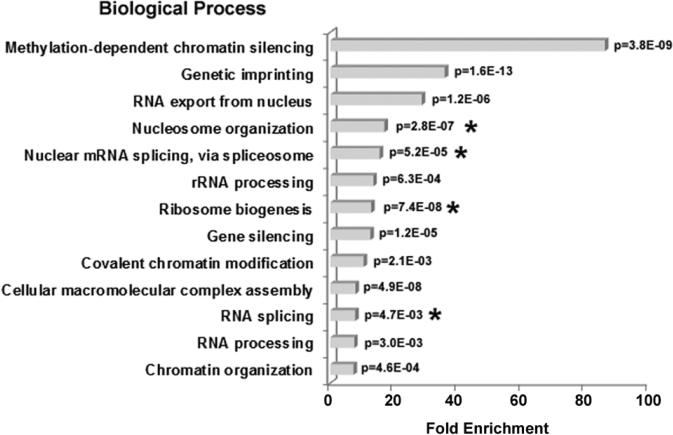
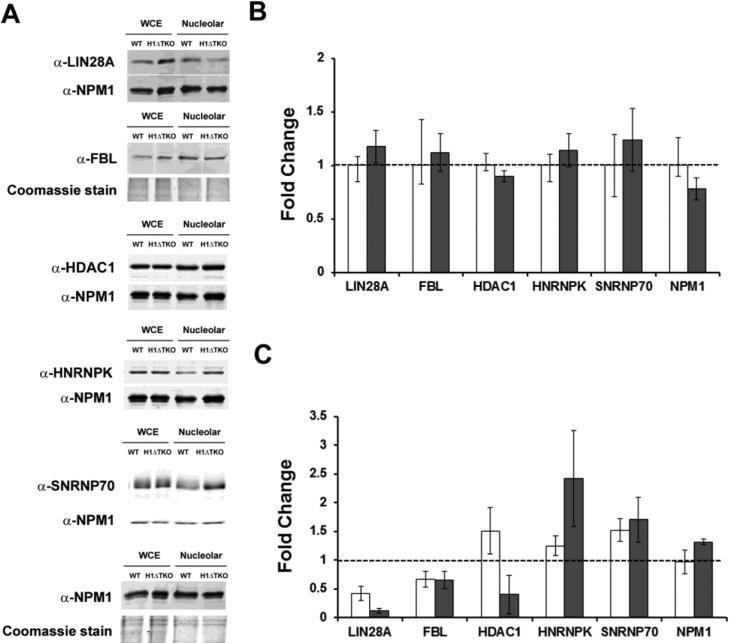
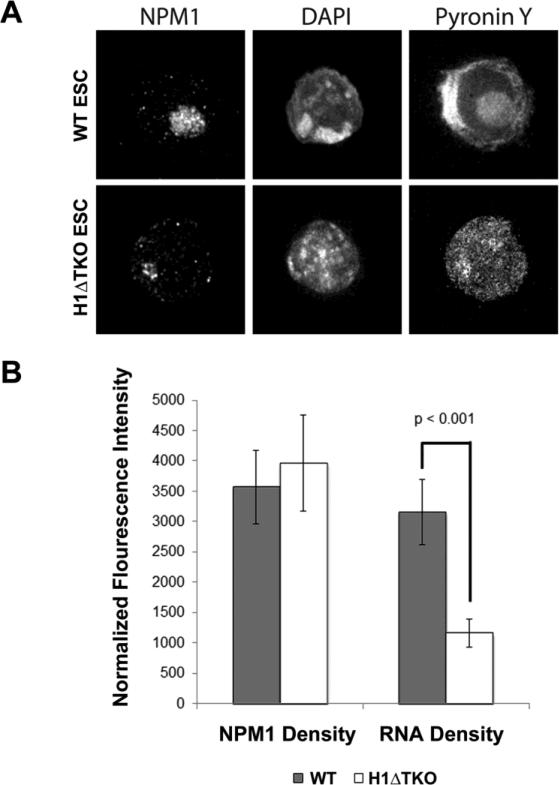
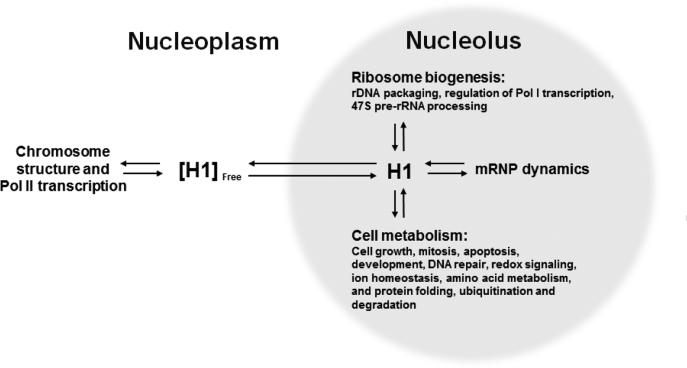
To investigate the relationship between linker histone H1 and protein-protein interactions in the nucleolus, we used biochemical and proteomics approaches to characterize nucleoli purified from cultured human and mouse cells. Mass spectrometry identified 175 proteins in human T cell nucleolar extracts that bound to Sepharose-immobilized H1 in vitro. Gene ontology analysis found significant enrichment for H1 binding proteins with functions related to nucleolar chromatin structure and RNA polymerase I transcription regulation, rRNA processing, and mRNA splicing. Consistent with the affinity binding results, H1 existed in large (400 to >650kDa) macromolecular complexes in human T cell nucleolar extracts. To complement the biochemical experiments, we investigated the effects of in vivo H1 depletion on protein content and structural integrity of the nucleolus using the H1 triple isoform knockout (H1ΔTKO) mouse embryonic stem cell (mESC) model system. Proteomic profiling of purified wild-type mESC nucleoli identified a total of 613 proteins, only ~60% of which were detected in the H1 mutant nucleoli. Within the affected group, spectral counting analysis quantitated 135 specific nucleolar proteins whose levels were significantly altered in H1ΔTKO mESC. Importantly, the functions of the affected proteins in mESC closely overlapped with those of the human T cell nucleolar H1 binding proteins. Immunofluorescence microscopy of intact H1ΔTKO mESC demonstrated both a loss of nucleolar RNA content and altered nucleolar morphology resulting from in vivo H1 depletion. We conclude that H1 organizes and maintains an extensive protein-protein interaction network in the nucleolus required for nucleolar structure and integrity.
Keywords: chromatin structure and gene expression; mass spectrometry; messenger RNA splicing; protein–protein interactions; ribosome biogenesis.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
References
-
- Happel N, Doenecke D. Histone H1 and its isoforms: contribution to chromatin structure and function. Gene. 2009;431:1–12. - PubMed
-
- Kowalski A, Palyga J. Linker histone subtypes and their allelic variants. Cell biology international. 2012;36:981–996. - PubMed
-
- Izzo A, Kamieniarz K, Schneider R. The histone H1 family: specific members, specific functions? Biological chemistry. 2008;389:333–343. - PubMed
-
- Luger K, Mader AW, Richmond RK, Sargent DF, Richmond TJ. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature. 1997;389:251–260. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
