

A DCM study of spectral asymmetries in feedforward and feedback connections between visual areas V1 and V4 in the monkey
- PMID: 25585017
- PMCID: PMC4334664
- DOI: 10.1016/j.neuroimage.2014.12.081
A DCM study of spectral asymmetries in feedforward and feedback connections between visual areas V1 and V4 in the monkey
Abstract
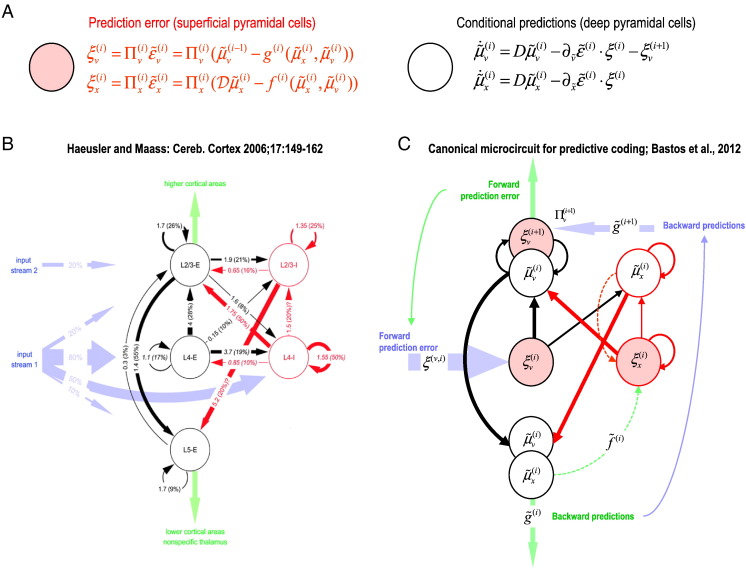
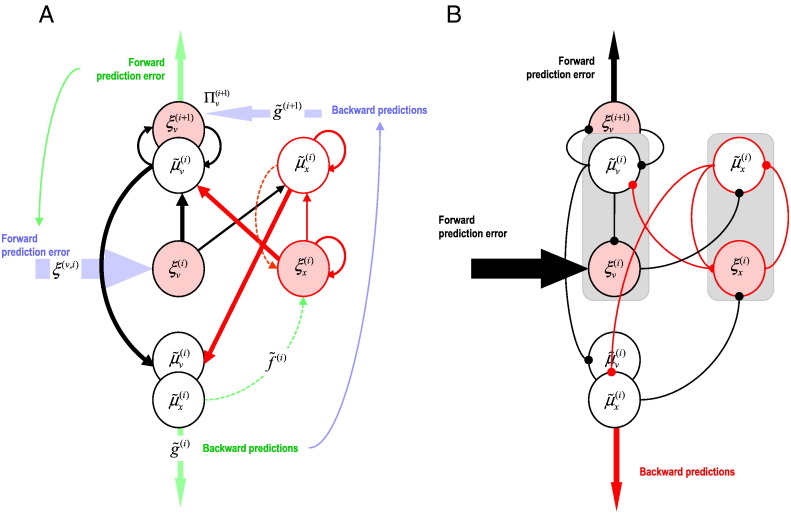
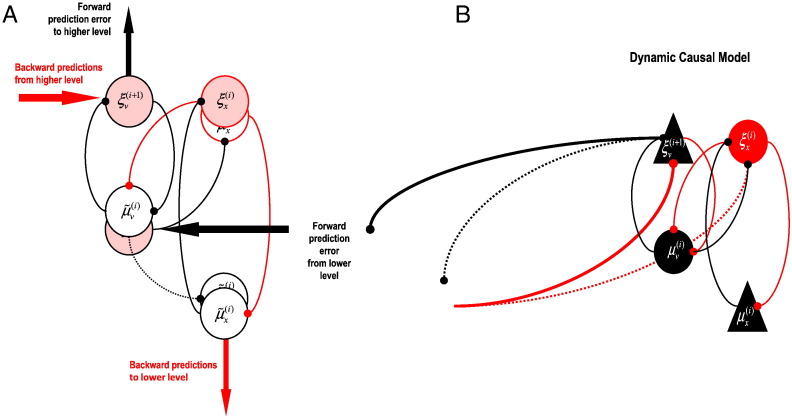
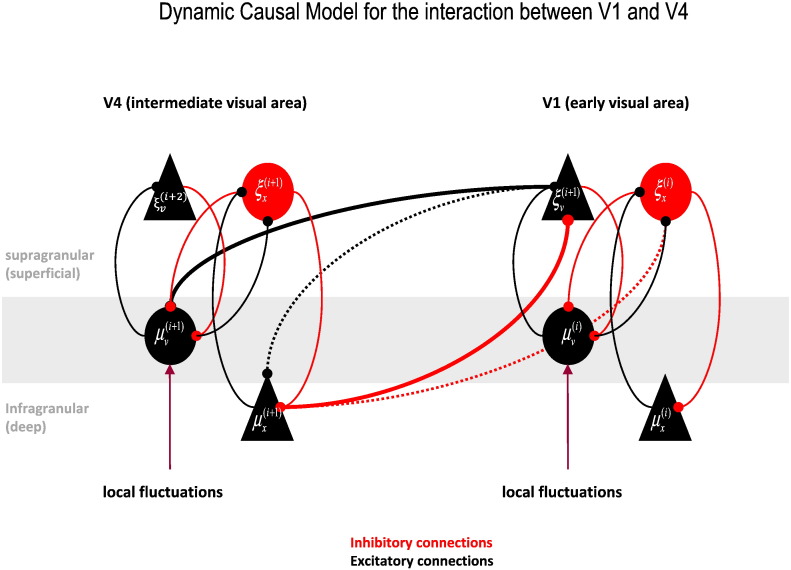
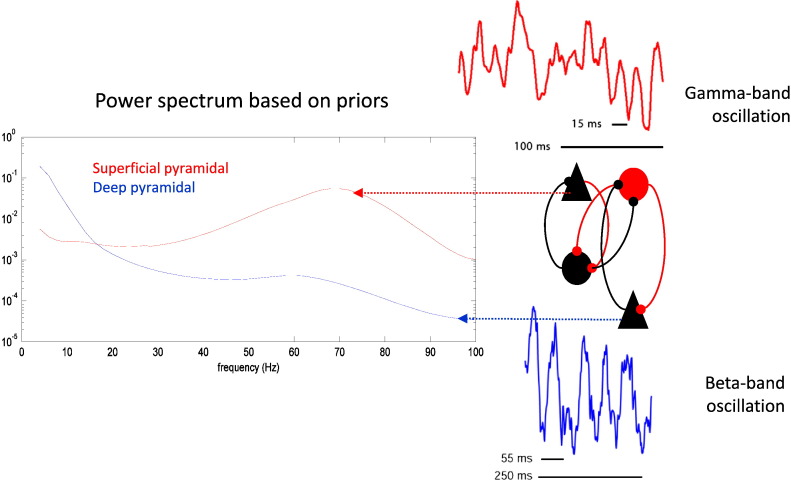
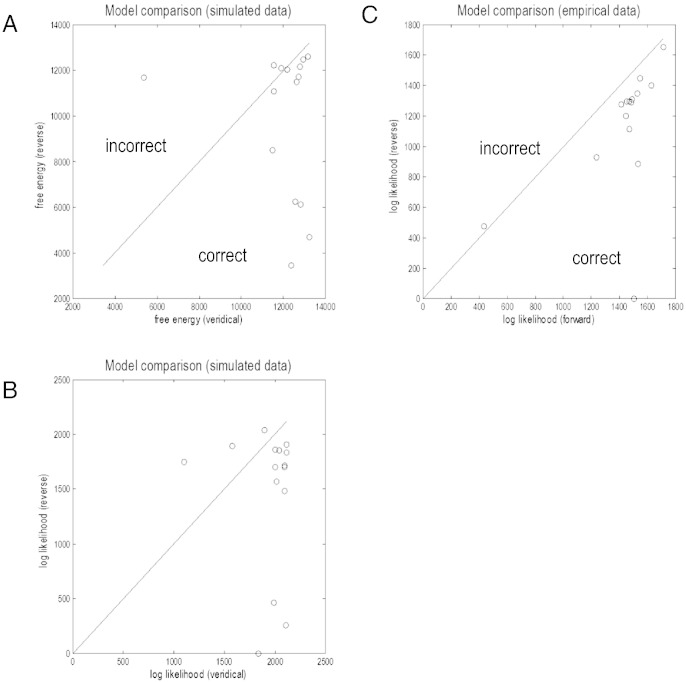
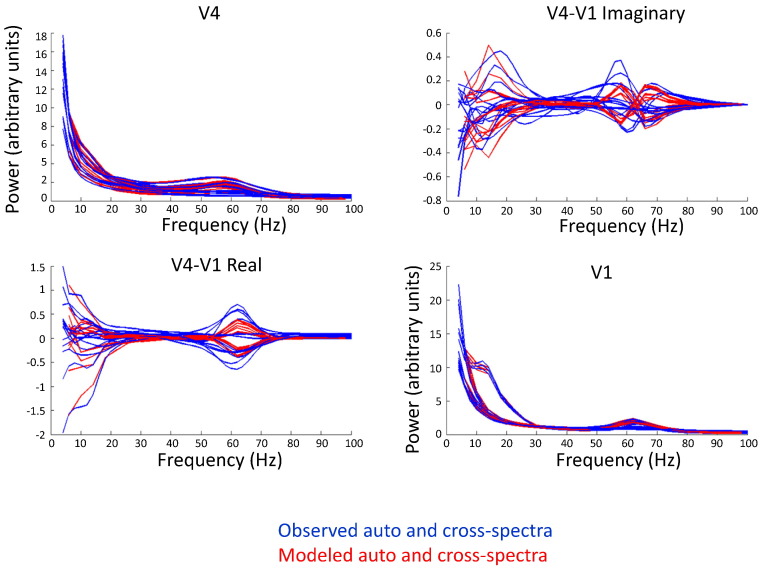
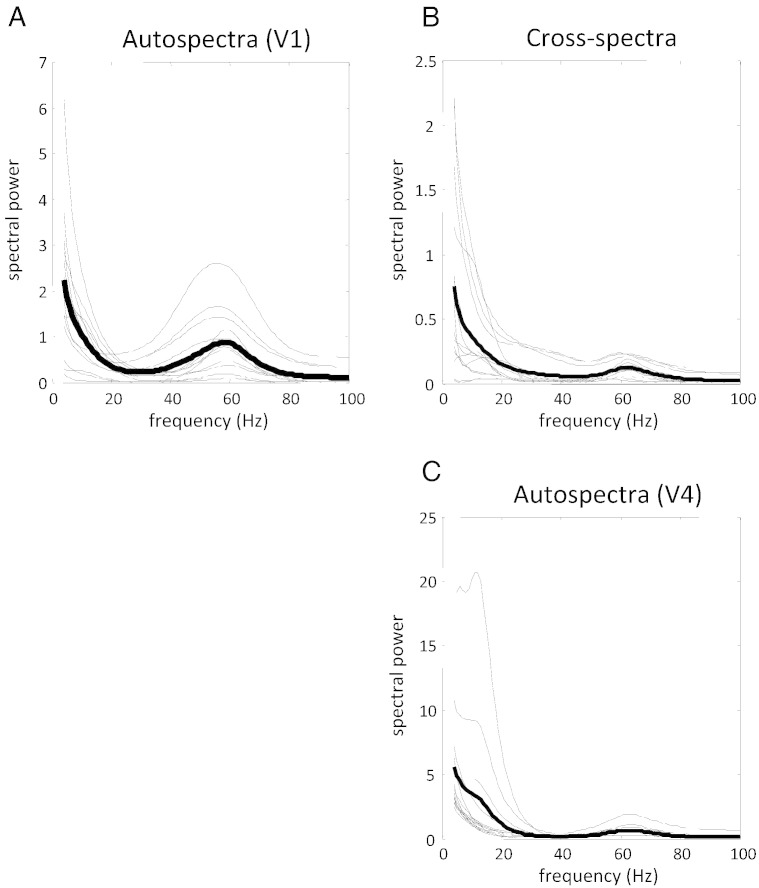
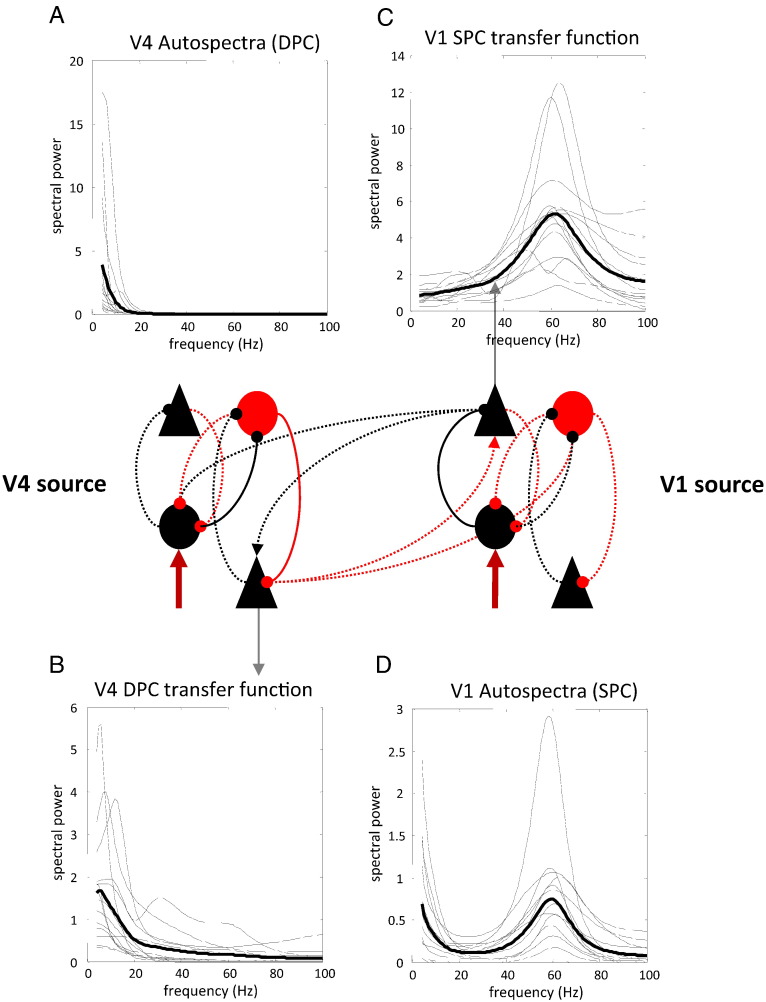
This paper reports a dynamic causal modeling study of electrocorticographic (ECoG) data that addresses functional asymmetries between forward and backward connections in the visual cortical hierarchy. Specifically, we ask whether forward connections employ gamma-band frequencies, while backward connections preferentially use lower (beta-band) frequencies. We addressed this question by modeling empirical cross spectra using a neural mass model equipped with superficial and deep pyramidal cell populations-that model the source of forward and backward connections, respectively. This enabled us to reconstruct the transfer functions and associated spectra of specific subpopulations within cortical sources. We first established that Bayesian model comparison was able to discriminate between forward and backward connections, defined in terms of their cells of origin. We then confirmed that model selection was able to identify extrastriate (V4) sources as being hierarchically higher than early visual (V1) sources. Finally, an examination of the auto spectra and transfer functions associated with superficial and deep pyramidal cells confirmed that forward connections employed predominantly higher (gamma) frequencies, while backward connections were mediated by lower (alpha/beta) frequencies. We discuss these findings in relation to current views about alpha, beta, and gamma oscillations and predictive coding in the brain.
Keywords: Beta oscillations; Computation; Connectivity; Dynamic causal modeling; Gamma oscillations; Neuronal; Synchronization coherence; Transfer functions.
Copyright © 2015. Published by Elsevier Inc.
Figures
References
-
- Arnal L.H., Wyart V., Giraud A.-L. Transitions in neural oscillations reflect prediction errors generated in audiovisual speech. Nat. Neurosci. 2011;14:797–801. - PubMed
-
- Bastos A., Bosman C., Schoffelen J.M., Oostenveld R., Fries Interareal directed interactions and their modulation by selective attention assessed with high density electrocorticography in monkey. Front. Hum. Neurosci. 2011;5
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
