

MET1 is a thylakoid-associated TPR protein involved in photosystem II supercomplex formation and repair in Arabidopsis
- PMID: 25587003
- PMCID: PMC4330576
- DOI: 10.1105/tpc.114.132787
MET1 is a thylakoid-associated TPR protein involved in photosystem II supercomplex formation and repair in Arabidopsis
Abstract
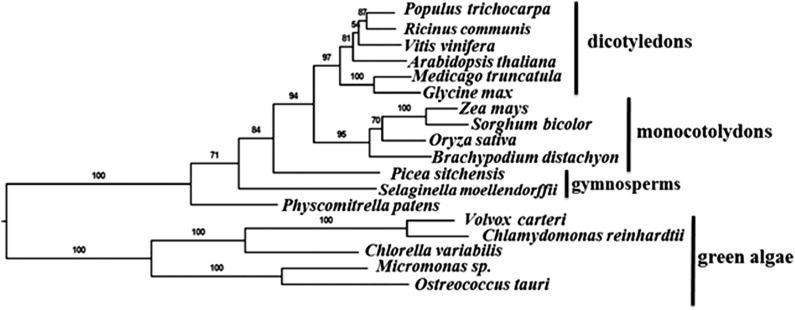
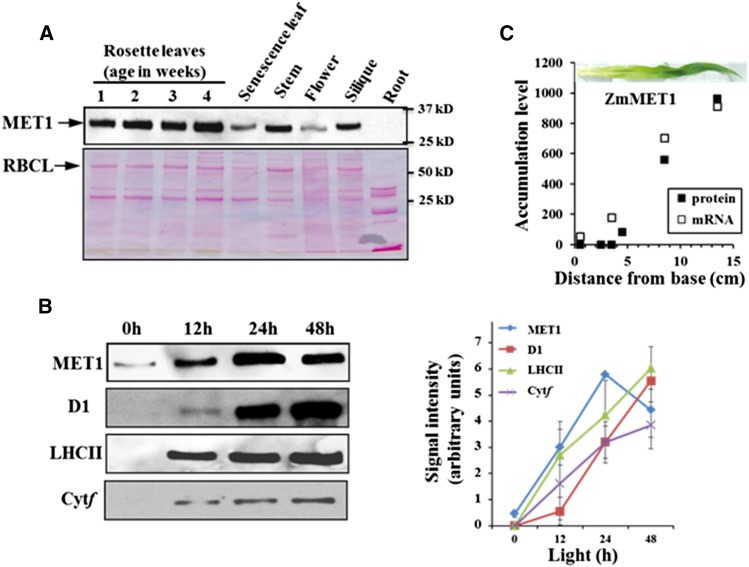
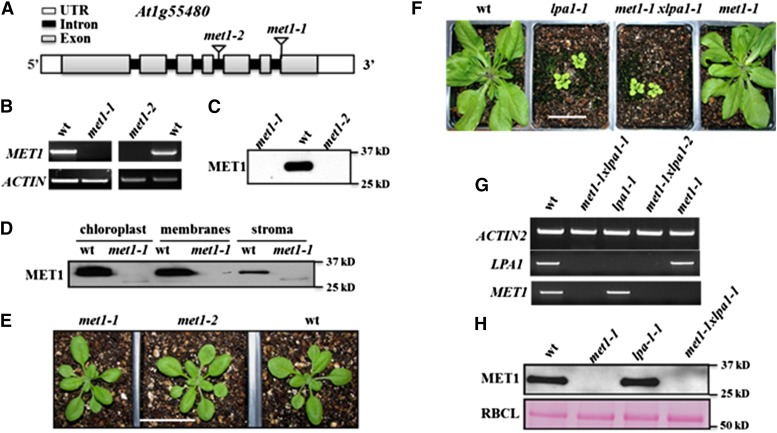
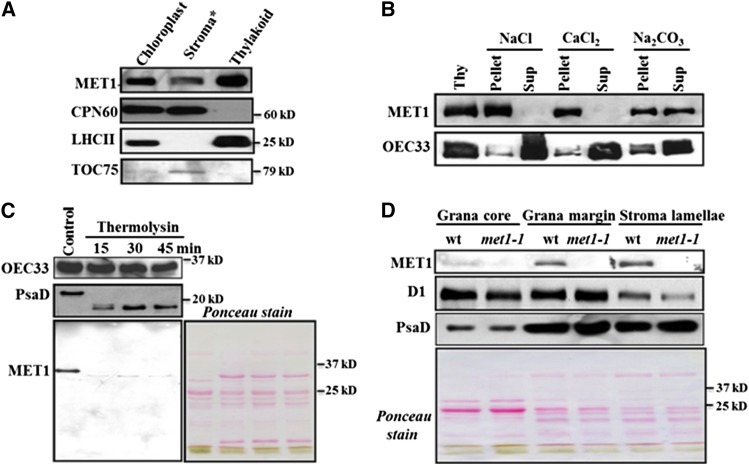
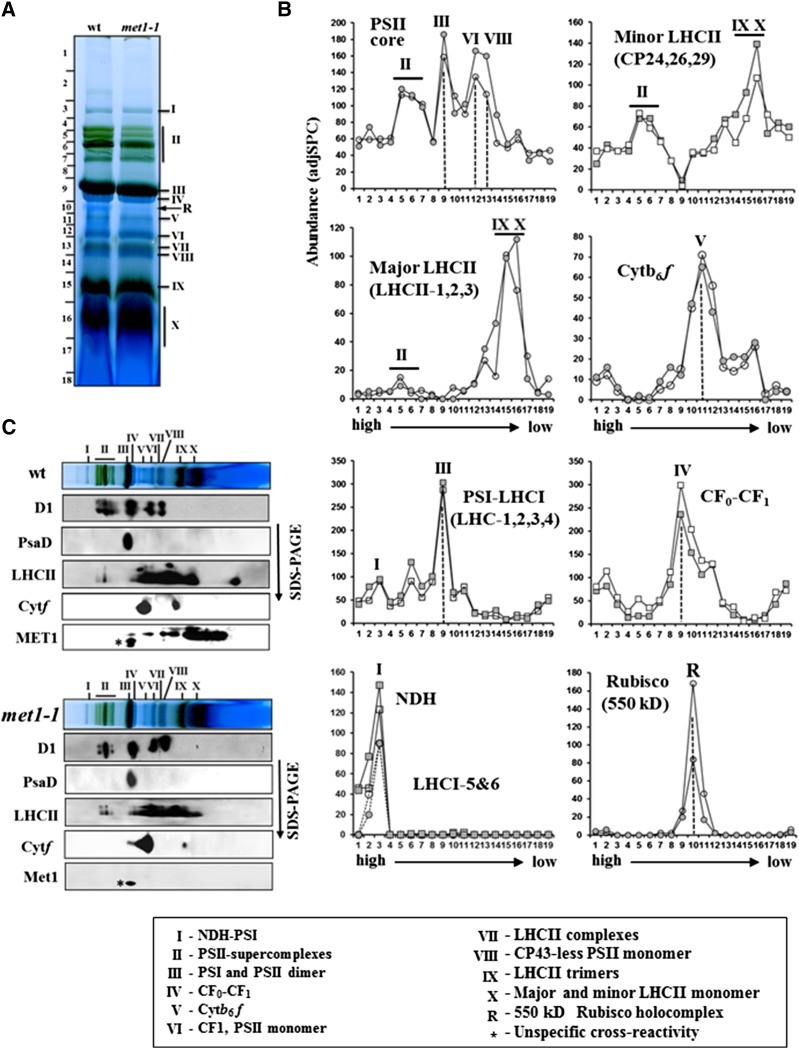
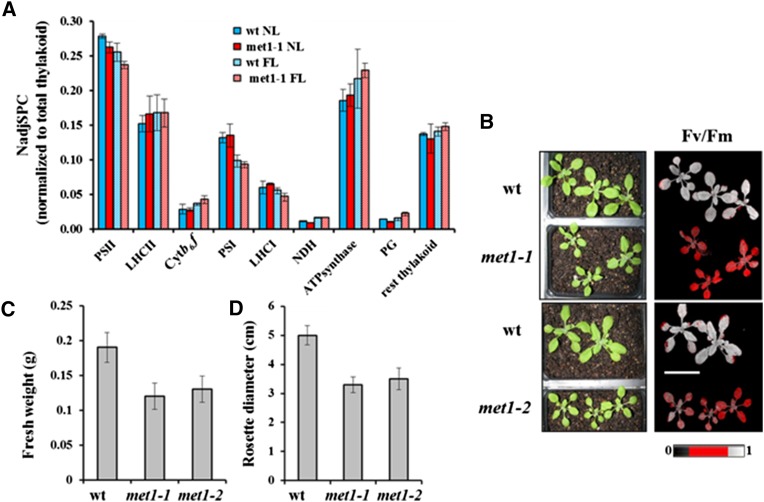
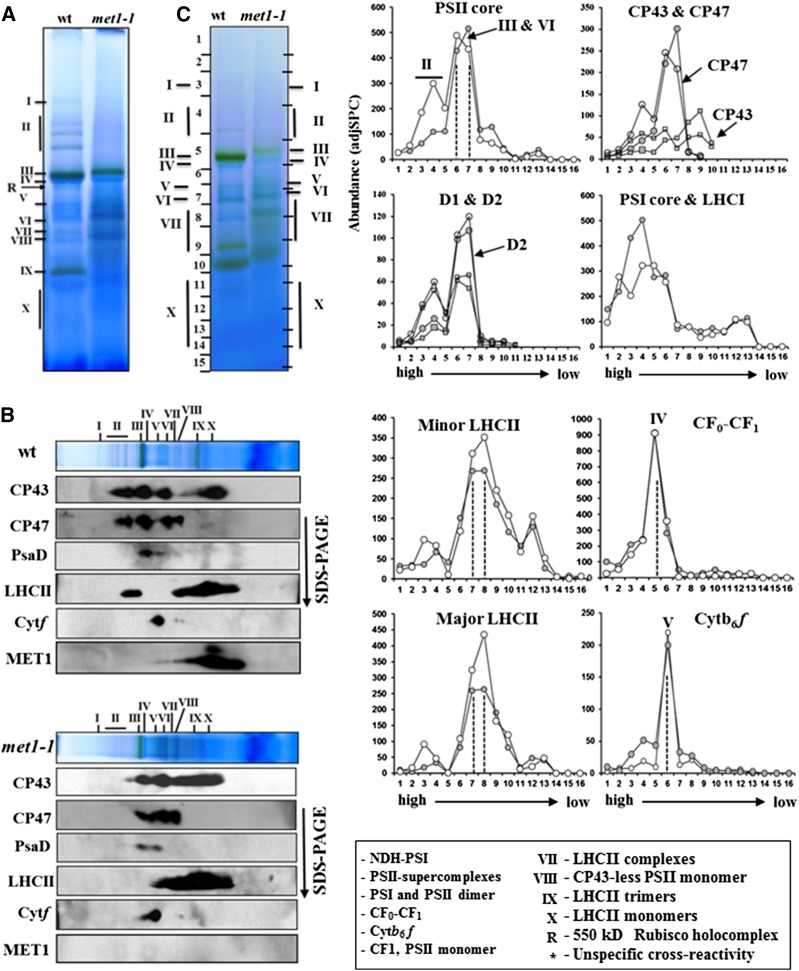
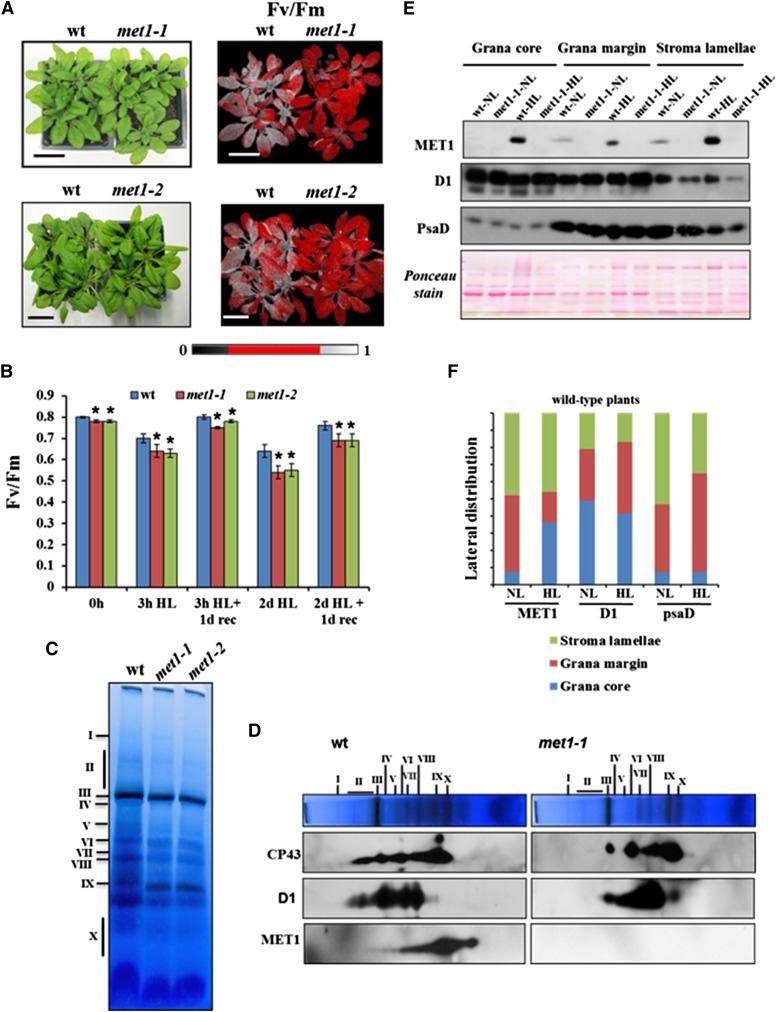
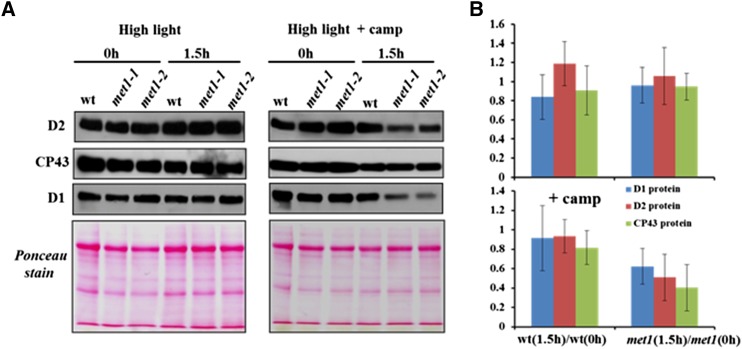
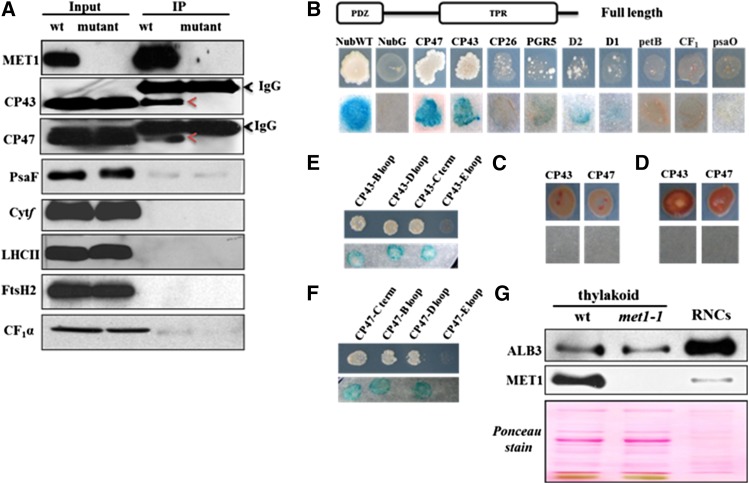
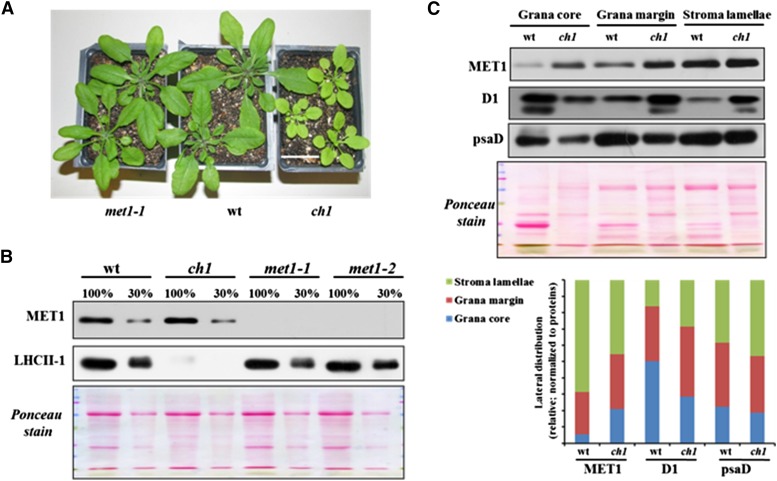
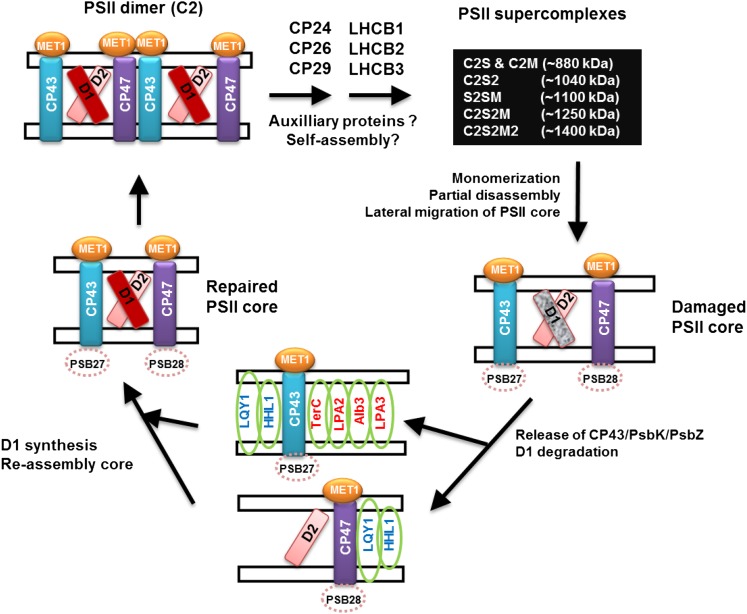
Photosystem II (PSII) requires constant disassembly and reassembly to accommodate replacement of the D1 protein. Here, we characterize Arabidopsis thaliana MET1, a PSII assembly factor with PDZ and TPR domains. The maize (Zea mays) MET1 homolog is enriched in mesophyll chloroplasts compared with bundle sheath chloroplasts, and MET1 mRNA and protein levels increase during leaf development concomitant with the thylakoid machinery. MET1 is conserved in C3 and C4 plants and green algae but is not found in prokaryotes. Arabidopsis MET1 is a peripheral thylakoid protein enriched in stroma lamellae and is also present in grana. Split-ubiquitin assays and coimmunoprecipitations showed interaction of MET1 with stromal loops of PSII core components CP43 and CP47. From native gels, we inferred that MET1 associates with PSII subcomplexes formed during the PSII repair cycle. When grown under fluctuating light intensities, the Arabidopsis MET1 null mutant (met1) showed conditional reduced growth, near complete blockage in PSII supercomplex formation, and concomitant increase of unassembled CP43. Growth of met1 in high light resulted in loss of PSII supercomplexes and accelerated D1 degradation. We propose that MET1 functions as a CP43/CP47 chaperone on the stromal side of the membrane during PSII assembly and repair. This function is consistent with the observed differential MET1 accumulation across dimorphic maize chloroplasts.
© 2015 American Society of Plant Biologists. All rights reserved.
Figures
References
-
- Aro E.M., Suorsa M., Rokka A., Allahverdiyeva Y., Paakkarinen V., Saleem A., Battchikova N., Rintamäki E. (2005). Dynamics of photosystem II: a proteomic approach to thylakoid protein complexes. J. Exp. Bot. 56: 347–356. - PubMed
-
- Ballottari M., Girardon J., Dall’osto L., Bassi R. (2012). Evolution and functional properties of photosystem II light harvesting complexes in eukaryotes. Biochim. Biophys. Acta 1817: 143–157. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
