

Characterization of chloroplast protein import without Tic56, a component of the 1-megadalton translocon at the inner envelope membrane of chloroplasts
- PMID: 25588737
- PMCID: PMC4348784
- DOI: 10.1104/pp.114.255562
Characterization of chloroplast protein import without Tic56, a component of the 1-megadalton translocon at the inner envelope membrane of chloroplasts
Abstract
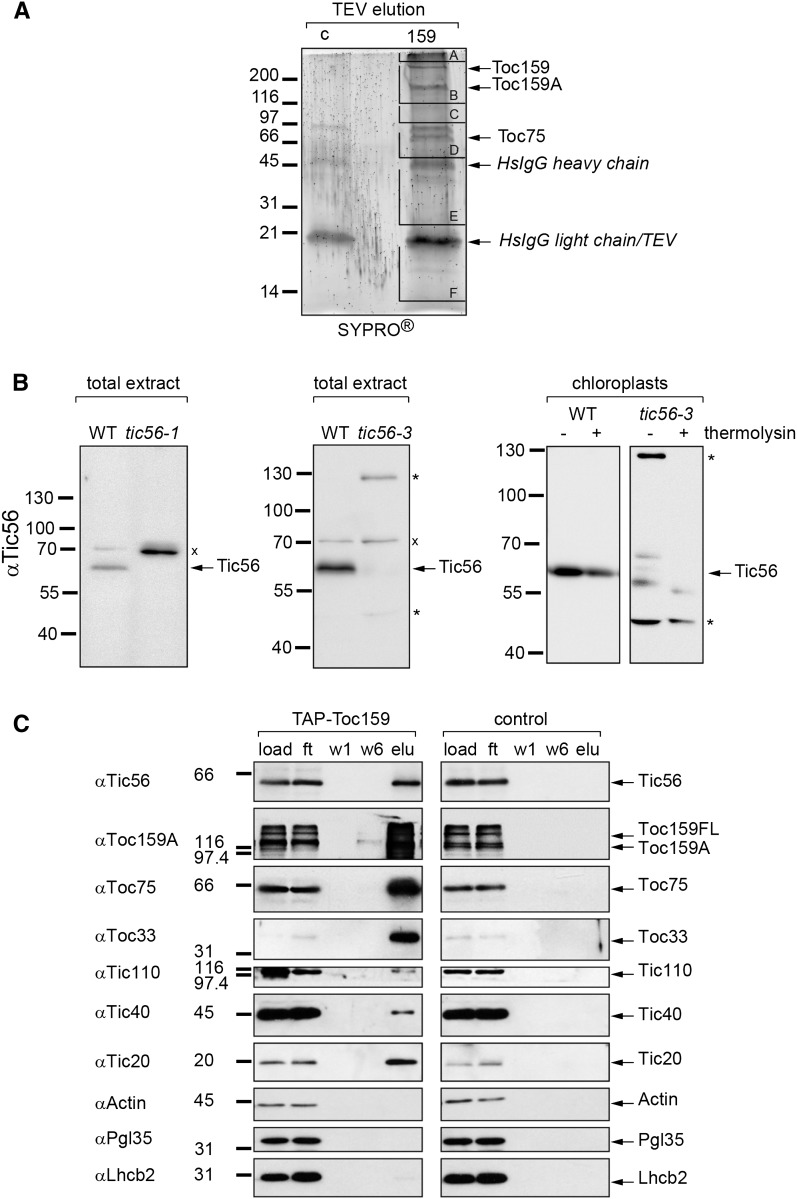
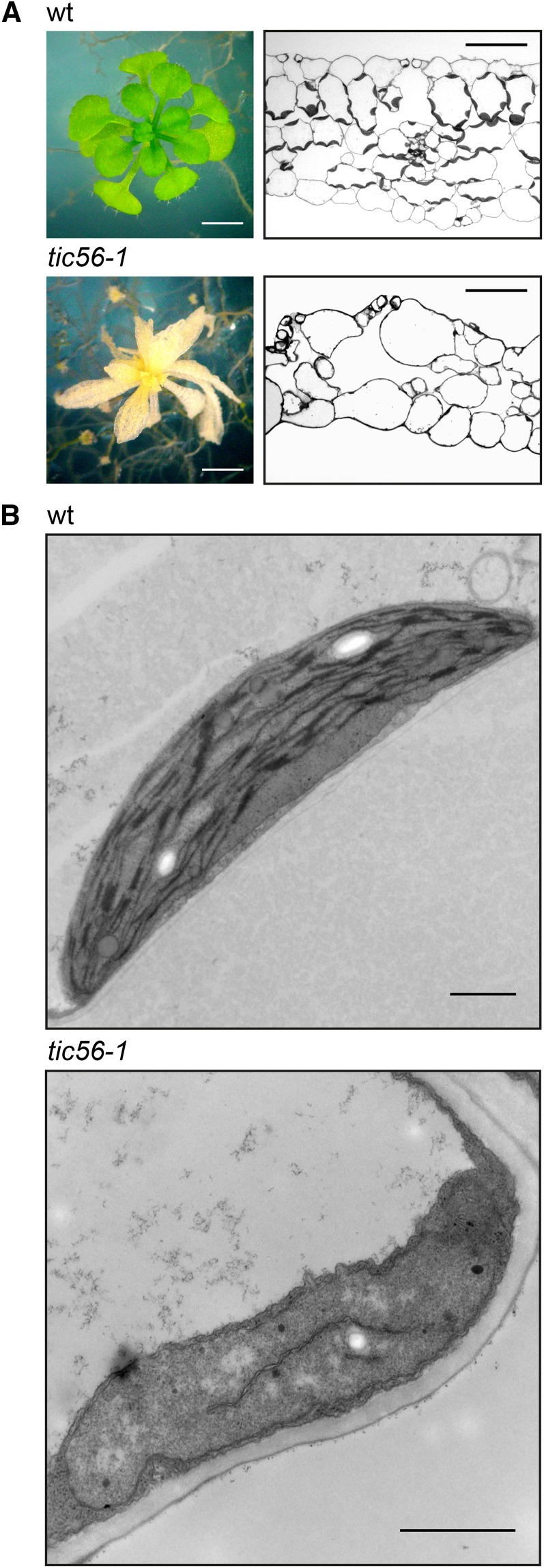
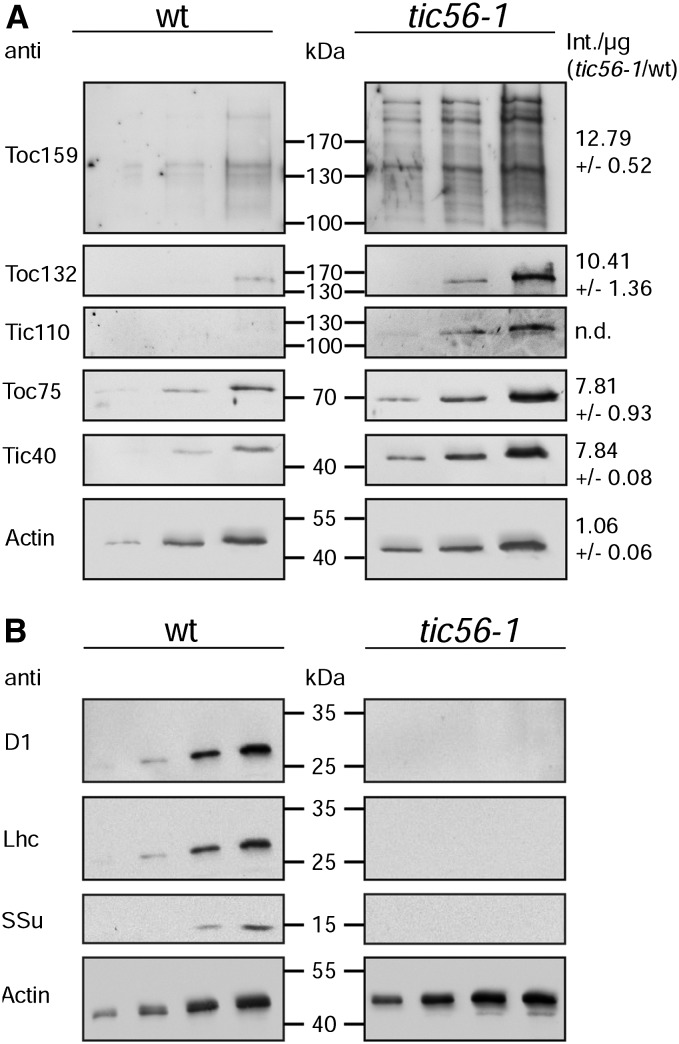
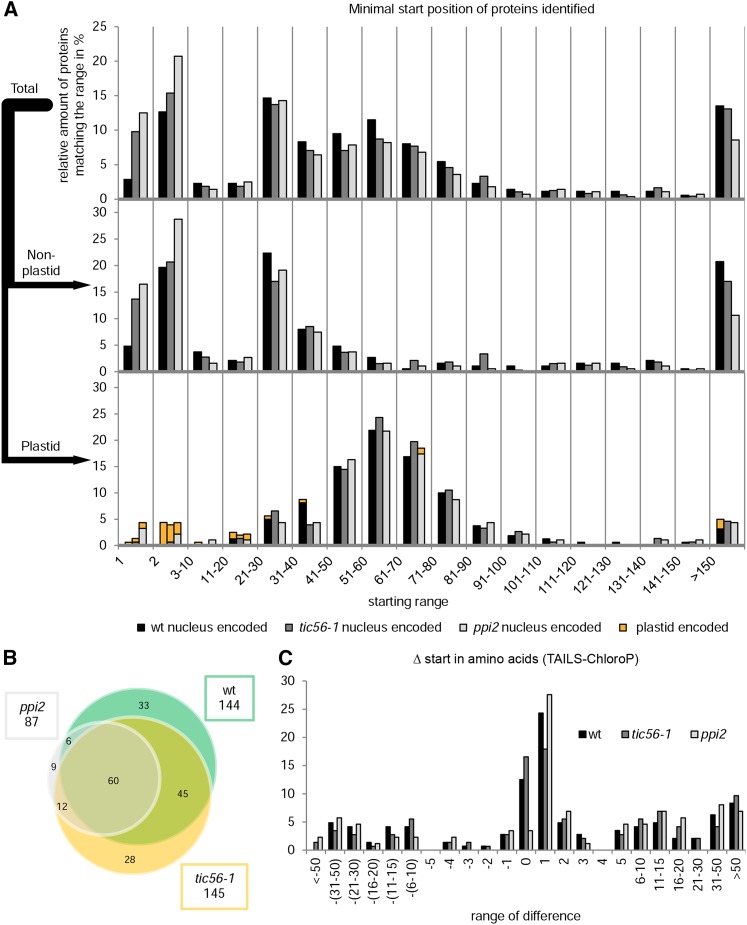
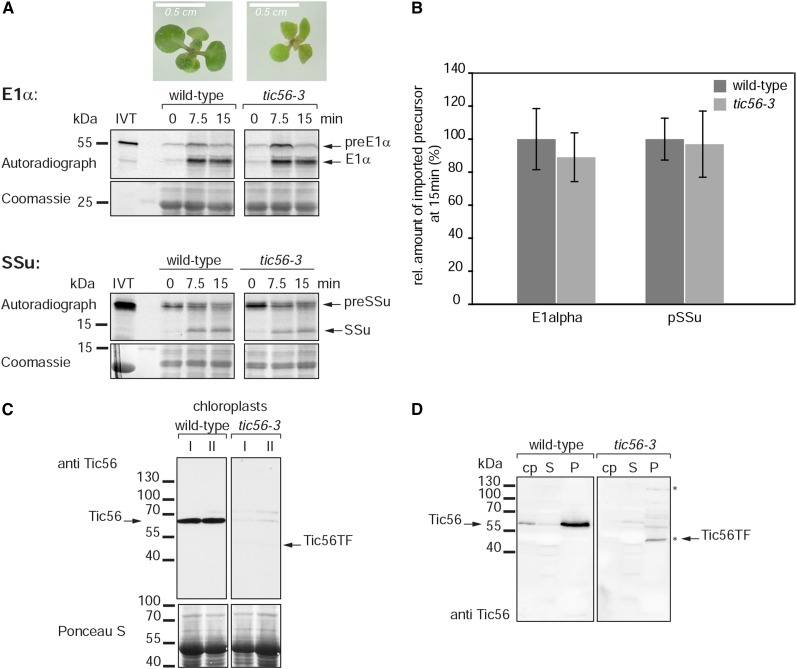
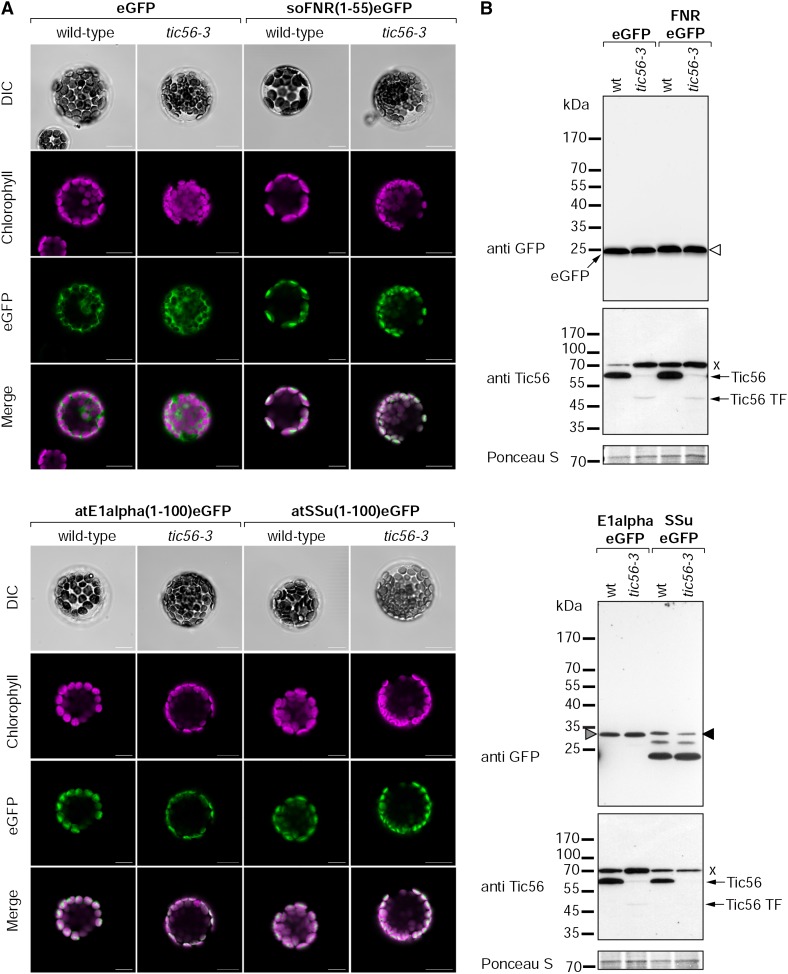
We report on the characterization of Tic56, a unique component of the recently identified 1-MD translocon at the inner envelope membrane of chloroplasts (TIC) in Arabidopsis (Arabidopsis thaliana) comprising Tic20, Tic100, and Tic214. We isolated Tic56 by copurification with Tandem Affinity Purification-tagged Toc159 in the absence of precursor protein, indicating spontaneous and translocation-independent formation of the translocon at the outer envelope membrane of chloroplasts (TOC) and TIC supercomplexes. Tic56 mutant plants have an albino phenotype and are unable to grow without an external carbon source. Using specific enrichment of protein amino termini, we analyzed the tic56-1 and plastid protein import2 (toc159) mutants to assess the in vivo import capacity of plastids in mutants of an outer and inner envelope component of the anticipated TOC-TIC supercomplex. Inboth mutants, we observed processing of several import substrates belonging to various pathways. Our results suggest that despite the severe developmental defects, protein import into Tic56-deficient plastids is functional to a considerable degree, indicating the existence of alternative translocases at the inner envelope membrane.
Figures
References
-
- Balsera M, Goetze TA, Kovács-Bogdán E, Schürmann P, Wagner R, Buchanan BB, Soll J, Bölter B (2009) Characterization of Tic110, a channel-forming protein at the inner envelope membrane of chloroplasts, unveils a response to Ca2+ and a stromal regulatory disulfide bridge. J Biol Chem 284: 2603–2616 - PubMed
-
- Bauer J, Chen K, Hiltbunner A, Wehrli E, Eugster M, Schnell D, Kessler F (2000) The major protein import receptor of plastids is essential for chloroplast biogenesis. Nature 403: 203–207 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
