

Functional characterization and developmental expression profiling of gibberellin signalling components in Vitis vinifera
- PMID: 25588745
- PMCID: PMC4339604
- DOI: 10.1093/jxb/eru504
Functional characterization and developmental expression profiling of gibberellin signalling components in Vitis vinifera
Abstract
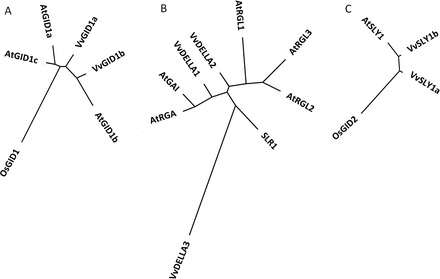
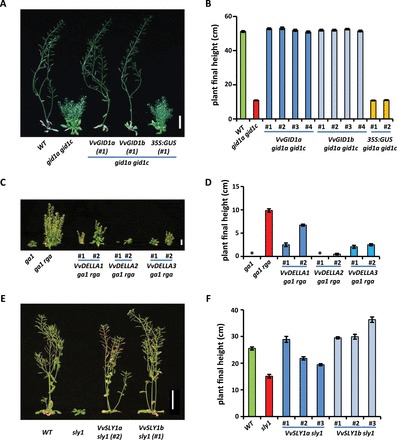
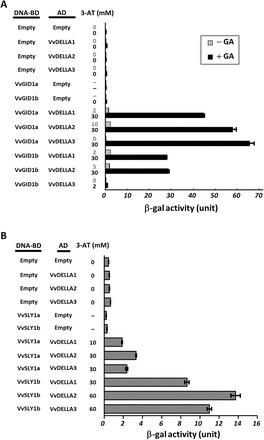
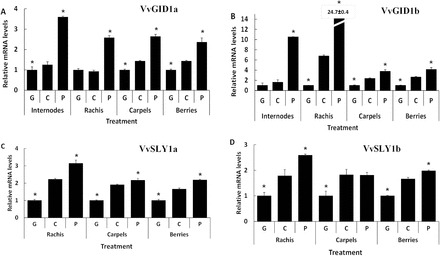
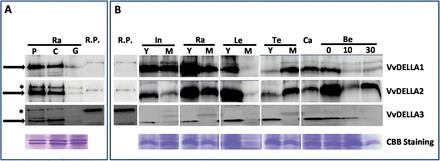
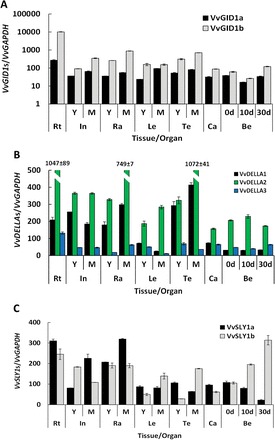
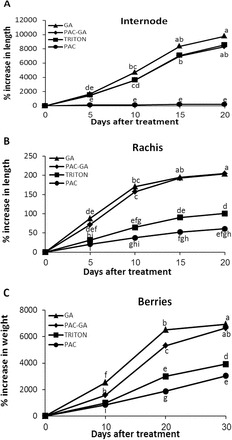
Gibberellins (GAs) regulate numerous developmental processes in grapevine (Vitis vinifera) such as rachis elongation, fruit set, and fruitlet abscission. The ability of GA to promote berry enlargement has led to its indispensable use in the sternospermocarpic ('seedless') table grape industry worldwide. However, apart from VvGAI1 (VvDELLA1), which regulates internode elongation and fruitfulness, but not berry size of seeded cultivars, little was known about GA signalling in grapevine. We have identified and characterized two additional DELLAs (VvDELLA2 and VvDELLA3), two GA receptors (VvGID1a and VvGID1b), and two GA-specific F-box proteins (VvSLY1a and VvSLY1b), in cv. Thompson seedless. With the exception of VvDELLA3-VvGID1b, all VvDELLAs interacted with the VvGID1s in a GA-dependent manner in yeast two-hybrid assays. Additionally, expression of these grape genes in corresponding Arabidopsis mutants confirmed their functions in planta. Spatiotemporal analysis of VvDELLAs showed that both VvDELLA1 and VvDELLA2 are abundant in most tissues, except in developing fruit where VvDELLA2 is uniquely expressed at high levels, suggesting a key role in fruit development. Our results further suggest that differential organ responses to exogenous GA depend on the levels of VvDELLA proteins and endogenous bioactive GAs. Understanding this interaction will allow better manipulation of GA signalling in grapevine.
Keywords: F-box proteins; GA receptors; VvDELLA proteins.; gibberellin signalling; gibberellins (GAs); grapevine (Vitis vinifera).
© The Author 2015. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
References
-
- Acheampong AK, Rotman A, Zaheng C, Keren A, Halaly T, Crane O, Ogrodovitch A, Or E. 2010. A method for isolating total RNA from mature buds and other woody tissues of Vitis Vinifera. In: Delrot S, Or E, Babaresco L, Grando S, eds. Methodologies and results in grapevine research. The Netherlands: Springer, 301–307.
-
- Agüero C, Vigliocco A, Abdala G, Tizio R. 2000. Effect of gibberellic acid and uniconazol on embryo abortion in the stenospermocarpic grape cultivars Emperatriz and Perlon. Plant Growth Regulation 30, 9–16.
-
- Ben-Tal Y. 1990. Effects of gibberellin treatments on ripening and berry drop from Thompson Seedless grapes. American Journal of Enology and Viticulture 41, 142–146.
-
- Boll S, Lange T, Hofmann H, Schwappach P. 2009. Correspondence between gibberellin-sensitivity and pollen tube abundance in different seeded vine varieties. Mitteilungen Klosterneuburg 59, 129–133.
-
- Boss PK, Thomas MR. 2002. Association of dwarfism and floral induction with a grape ‘green revolution’ mutation. Nature 416, 847–850. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
